Warning: Trying to access array offset on value of type null in /home/elisew5/public_html/drpeterwitt.com/wp-content/plugins/contextual-related-posts/includes/content.php on line 49
Images Collection
View this article in Search Friendly Plain Text
NOTE: This plain text article interpretation has been digitally created by OCR software to estimate the article text, to help both users and search engines find relevant article content. To read the actual article text, view or download the PDF above.
484
The American Midland Naturalist
94(2)
Change of Web Site in Argiope Spiders (Araneidae)
Abstract: Argiope aurantia and A. trifasciata are more likely to change web site within 3 days after molting than in the days before molting, both in the laboratory and in the field, and earlier instars of A. aurantia are more likely to change web site than are later instars. Both species show web-site tenacities above 80% in the field.
Introduction
Since most animals have higher fitness in certain places, behavior has evolved which results in the selection of particular habitats (Levins, 1968). Many animals simply reduce the rate of travel in preferred localities (Sale, 1969; Klopfer, 1965; Wecker, 1963).
The web of spiders serves both as the trap for food and as the spider’s living quarters. The building of the web itself has been studied (Witt et al., 1968; Reed et al., 1969), but the behavior which relates the web to the site has not. “Web-site tenacity” can be defined as the percentage probability that, once it has built a web, the spider will remain at the same web site from one day to the next ‘(Enders, 1973). Here I describe internal changes which affect the web-site tenacity in two species of spiders in the genus Argiope in the family Araneidae. This paper is the first formal study of web-site tenacity as it relates to selection of a place to live.
Observations in the Field MethodsU-Argiope aurantia (Lucas) and A. trifasciata (Forskal) are large orb web spiders which are abundant in the United States. Observations of these species were made during the daytime along the edges of road-cuts in Raleigh, North Carolina. Observations were made in such a way as to1 avoid gross disturbance either to the vegetation or to the web or web site, the vegetation to which the web was actually attached, The study areas were covered with Sericea Lespedeza (Lespedeza cuneata), a tall perennial herb which dies back to the ground each winter, and a considerable admixture of other plant species (Enders, 1974).
The instar of Argiope aurantia and of A. trifasciata spiders was estimated by comparison with preserved specimens of A. aurantia which had been reared in the laboratory. Second-instar spiders (Kaston, 1948) are those which emerge from the egg sac and are in the first web-building stage. One A. aurantia in the field had to be scored as the 12 th instar, but most females matured and stopped molting in instar nine or 10.
While still on the webs, spiders were marked on their dorsal body surface with individual patterns of rapidly drying paint. To identify the location of of each web, the supporting vegetation was marked, using masking tape. On successive days, I searched the areas where spiders had been marked. Each day I recorded whether the spiders remained at their old sites, whether they had molted, and whether any previously marked spiders could be found at new web sites. As marked animals disappeared, additional spiders in the vicinity were marked for observation, and animals were re-marked immediately after discovering that they had molted.
Web-site tenacity is equal to the number of spiders remaining at old sites divided by the sum of those that (a) remained at the old site, (b) were found elsewhere than the previous site, and (c) could neither themselves nor any part of the orb section of the old web be found (Table 1). Spiders which spontaneously left a web site removed the orb section before leaving. Observations of the webs of spiders known to be dead support the assumption that spiders have
1975
Notes and Discussion
485
died if their previous web can be relocated with no spider present. Thus, the denominator for the calculation of web-site tenacity is the number of spiders known to be marked the previous day less those which died (D in Table 1).
In certain cases another spider was observed in a web site when the original occupant of that web site had disappeared or changed web site. Ordinarily, the new occupant was a noticeably larger animal and sometimes a marked spider. The move by the original occupant on that day was excluded from consideration, because, on occasion, I have seen active intrusion onto a spider’s web by another (Enders, 1974). The move by the original occupant can thus be considered to have been forced. Blanke (1972) reports similar interactions in Cyrtophora citricola Forskal (Araneidae). Since fewer than 10 of my observations are excluded for this reason, such exclusion does not materially change the calculated web-site tenacity.
Most changes of web site were more than a diameter of the web and most were horizontal displacements. Criterion for change of web site was a minimum 10-cm change in the location of the hub of the web in any direction, including the vertical. Difficulty in deciding whether a change of web site had occurred was encountered only in observations made of smaller spiders, in which cases some shifts of less than 10 cm were noted, but were scored as an animal remaining at same site. Tests of significance were made by the use of chi-square tests not corrected for continuity (Snedecor and Cochran, 1967), the probability of 0.05 being used as the minimum level for reporting statistical significance.
Results.—The method of marking individuals caused much mortality of the tiny second and third instars of A. aurantia. But unmarked animals (observed as discussed under laboratory results) of these instars showed a web-site tenacity of roughly 50%, lower than that of larger spiders. Because it was impossible to avoid stepping on the vegetation near the web (Lowrie, 1963), and since second instars of this species leave sparse or short vegetation (Enders, 1973), those results might be discounted, but they fit with laboratory results discussed below.
Mortality was lower for larger araneids (Enders, 1974), such as 4% per day, one observation of 23 in category D (Table 1). Table 1 also shows the lower apparent mortality and movement rates of marked spiders, compared to unmarked ones, although the chi-square value is not statistically significant. Higher mortality and rate of movement had been considered possible effects of marking. A similar statistical analysis using unmarked Argiope spiders at later stages of the life history showed no significant differences between unmarked and marked individuals in observed web-site tenacity.
There were no gross differences in web-site tenacity among months and between species (Table 2). The lower web-site tenacity of second instar A. aurantia described above is reflected in Table 2 in the lower web-site tenacity for June, in which the model instar was the fifth instar.
Table 1.—Comparison of marked and unmarked A. aurantia observed on 4 successive days, 21-25 June 1970. Web-site tenacity = A / (A + B+C)
| Record on next day | No. of observations of unmarked spiders | No. of observations of marked spiders |
| A. Remained at site
B. & C. Found elsewhere or neither |
10 | 18 |
| web nor spider found | 6 | 4 |
| D. Old web found, but not spider | 1 | 1 |
| Calculated web-site tenacity | 62.5% | 81.8% |
486
The American Midland Naturalist
94(2)
Observations in the Laboratory
Methods.—-Web-site tenacity of early instars was observed in a 10-gal glass terrarium with a glass cover. This provided sufficient space for normal web building and for change of web site. Webs of these particular spiders were generally less than 10 cm in diam. Use of the terrarium avoided problems of gross disturbance and of disappearance of the small 1-mm-long second instar A. aurantia which had occurred in the field. Spiders were given water every day. Eye gnats {Hippelâtes sp.) were added to the terrarium once a week as food. Branches and wire had initially been placed in the terrarium for the spiders’ web building, and I did not destroy the webs during the period of observation.
These unmarked individuals of A. aurantia could often be identified due to (a) size differences among caged animals, (b) the small number of individuals and (c) the failure of other individuals of a similar size to change web site. These spiders originated from two cocoons. In larger cardboard and screen wire cages in the laboratory, which were at least five times the diameter of the webs, larger spiders were similarly given a choice of changing web site or remaining at the same one.
Results.—Many young spiders renewed their webs only every 2nd day. This occurred primarily in the second and third instars and was also observed in the field. In contrast, larger araneid spiders generally renewed their webs every night, as originally reported by Breed et al. (1964).
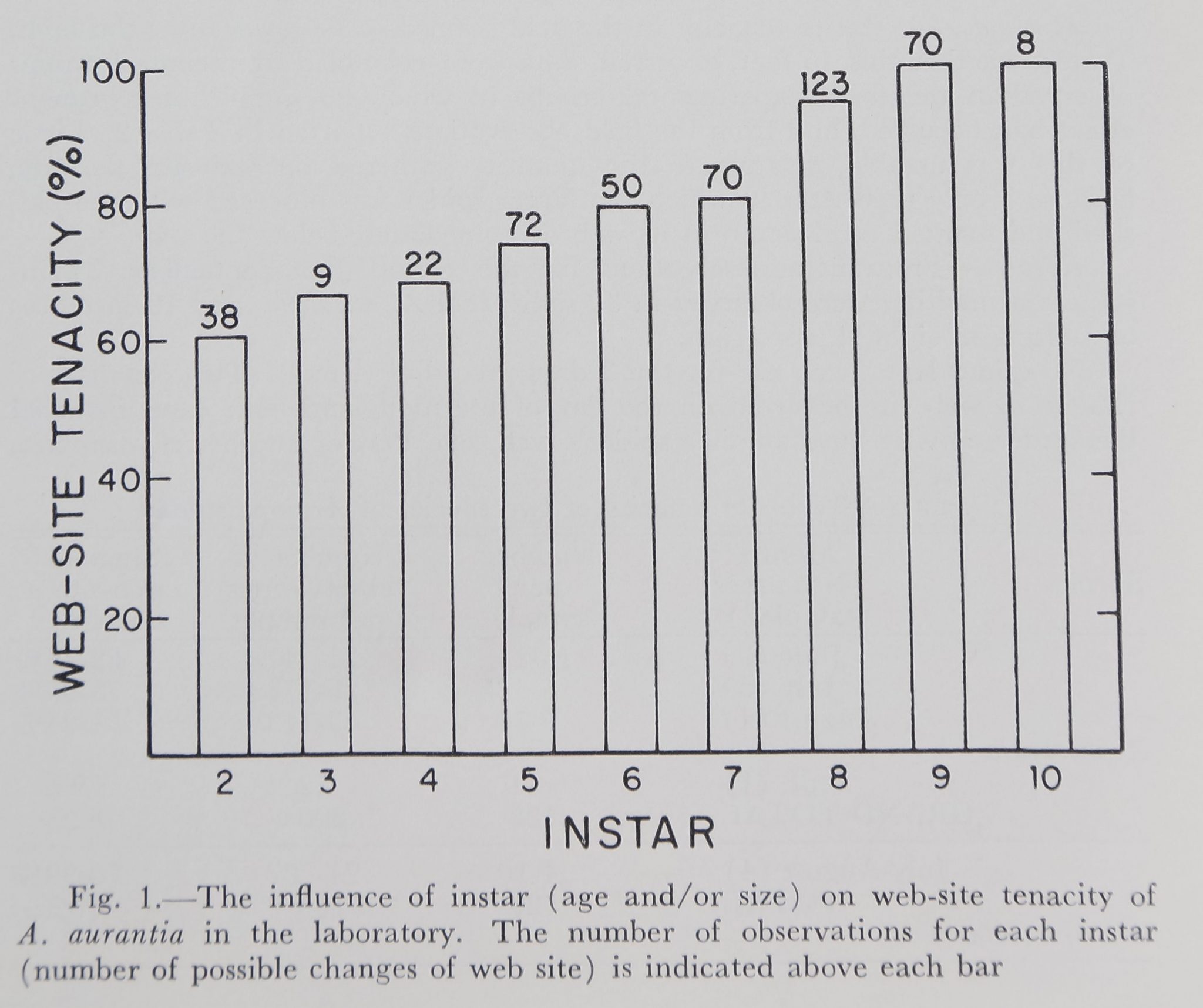
There was a marked and consistent increase in web-site tenacity with instar (age and/or size) of Argiope aurantia (Fig. 1).
Influence of Molting on Web-Site Tenacity
Methods.-^Web-site: tenacity in the field seemed to be lower after the molt. To test whether this, in fact, occurred, data were assembled from control groups observed in the field, experimental groups in which no significant treatment effect had occurred, and from the field observations reported in Table 2. Little of this was useable, relative to the quantity gathered on web-site tenacity, because I only used data in which the larger spider was observed with the old, shed and marked exoskeleton in its web or immediately below the web.
Results.—From field observations for the period June-September, 31 instances of molting were observed in 30 individual A. aurantia, and 10 instances of molting in eight A. trifasciata.
No spider left a web site for the 3 days preceding a molt. Two instances of change of web site occurred on the day of the molt, and both cases involved the spider moving onto another spider’s web, one that of another A. aurantia,
Table 2.—Web-site tenacity of two species of Argiope spider
| Month | Number | Number of | Range of |
| Species (No. areas | per | observations | web-site |
| sampled) | sample | per sample | tenacity |
| June (2) | 16-21 | 21- 41 | 62-76% |
| July (5) | 6-48 | 26-173 | 75-96% |
| August (4) | 9-14 | 43-113 | 81-93% |
| A. aurantia | |||
| Sept. (1) | 11 | 52 | 79% |
| GRAND TOTAL | 188 | 800 | 82% |
| July-August (4) | 4-10 | 21- 87 | 70-99% |
| TOTAL | 29 | 166 | 90% |
A. trifasciata
1975
Notes and Discussion
487
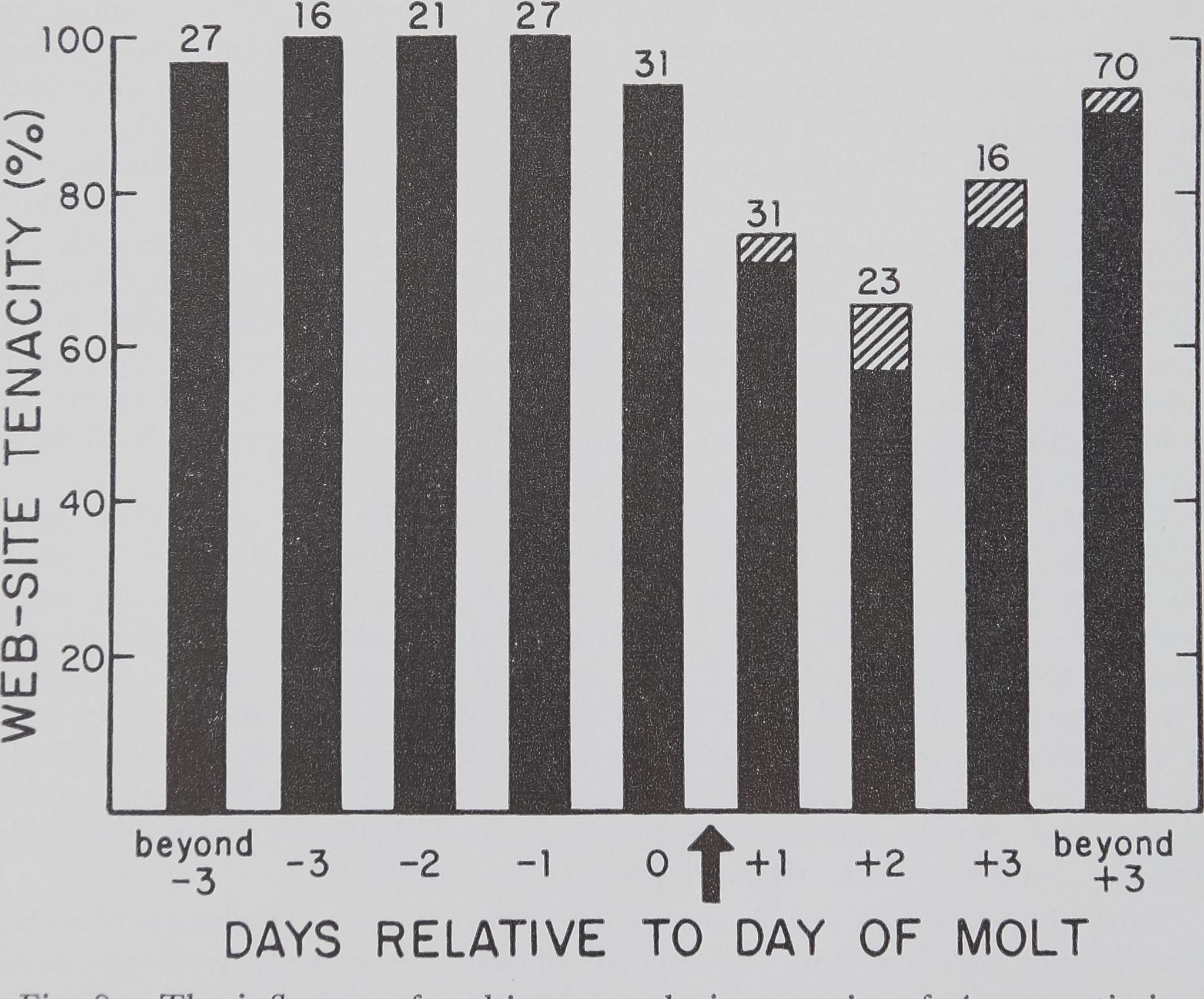
and the other that of an A. trifasciata. [These two web invasions are notable because web invasion occurred only in a very small percentage of cases (0.6%) of the entire set of 800 observations (Enders, 1974).] After the molt, A. aurantia spiders tended to change web site more often than before the molt. The difference between web-site tenacity up to the day of the molt and that to the day after the molt is shown in Figure 2. This difference is statistically significant (chi-square, 0.05 level) and web-site tenacity remained significantly lower for the 1st 3 days after the molt, compared to the period up to the day of the molt. Small movements of web position, less than the 10-cm criterion, commonly occurred only after the molt. Inclusion of such shifts of web position as changes of web site would emphasize the reduction in web-site tenacity occurring after the molt.
Argiope trifasciata showed a pattern of web-site tenacity similar to that shown in Figure 2. Numbering days as in Figure 2, no changes of web site occurred in 62 observations of A. trifasciata up to the day of the molt; two of 10 changed to day 1, and one of seven to day 2. No changes of web site were recorded in 10 observations made from day 2 to day 3 and beyond. Thus, the pattern of premolt web-site tenacity and a decline of web-site tenacity for the first few days after the molt was evident in A. trifasciata as well as A. aurantia.
In the laboratory, when A. aurantia were free to change web site, the difference between premolt and postmolt web-site tenacities was less marked than in the field—93% of 46 observations for 4 days before the molt contrasted to 81% for 36 observations for 3 days after the molt. A one-tailed (since anticipated from field data) chi-square test demonstrated a significant difference

488 The American Midland Naturalist 94(2)
between premolt and postmolt web-site tenacities. Laboratory data for A. trifasciata were not available.
Discussion
Mature male spiders leave their webs and locate females (Kaston, 1948). With this exception, web spiders seem to be more sedentary as adults than as immatures. Nevertheless, some change of web site has been observed even for adult females of the following species: (a) Psilochorus species (Pholcidae) (M. H. Muma, pers. comm.), (b) Uloborus diversus (Uloboridae) (Eberhard, 1971), (c) Achaearanea tepidariorum (Theridiidae) (Turnbull, 1964) and (d) several species in the Araneidae (Blanke, 1972; Argiope species, this study). Even though Eberhard (1971) marked webs and not web sites, his data do show if a web site has been abandoned. Based on web-site desertion rate (which includes mortality, unlike my calculations, above), his data therefore show U. diversus with a minimum web-site tenacity (100% minus website desertion rate) of 83%. Similarly, Blanke’s fragmentary data allow no precise calculation but show some web invasion and roughly 80% web-site tenacity in Cyrtophora citric ola. Thus, these authors found web-site tenacity to be at a level similar to that seen in Argiope species.
The Uloborus species (also Pholcidae and Theridiidae) generally do not rebuild the catching part of the web every day, unlike araneids. Little signifi-

Fig. 2.—The influence of molting on web-site tenacity of A. aurantia in the field. The arrow indicates the building of the fist web after the molt. The solid bars are calculations including shifts of web position of less than 10 cm as the web moved. The tops of the diagonally shaded portions of the bars show the level of web-site tenacity when 10-cm changes of web position are considered as no move of the web, as elsewhere in this paper
1975
Notes and Discussion
489
cance can therefore be placed upon the fact that Eberhard (1971) found older, larger U. diver sus disappeared from web sites less often than early instars (a maximum of 15% of last two instars, maximum of 13% for middle instars, and 18% for early instars, according to my recalculations from web-site desertion rates).
While Turnbull (1964) described a very strong positive effect of prey capture upon web-site tenacity in Achaearanea tepidariorum (Theridiidae), Enders (1972) found prey capture has little or no effect on web-site tenacity in Argiope aurantia, while Eberhard (1971) actually found a negative effect in Uloborus diver sus. Presumably, the Argiope and Uloborus species are not normally limited by prey abundance, so that they have not evolved any strong positive behavioral response to capture of prey.
A definite change in web-site tenacity with age and/or size in Argiope aurantia is reported here. Despite the fact that young spiders renewed their webs less often, thus giving less “opportunity” to leave a web site, the young left web sites more often. This change of web-site tenacity appears to be controlled by the onset of sexual maturity rather than by age or size per se. Since different individuals and the two sexes mature at different sizes, additional close observation of penultimate instar spiders might prove this hypothesis.
The significant reduction in web-site tenacity after molting is almost certainly related to the hormonal changes during that period. Krishnakumaran and Schneiderman (1970) have already demonstrated that topical insect ecdysone can cause a behavioral effect in an araneid spider, e.g,} the spinning of a strong retreat in which to molt by Araneus cornutus. Increased web-site tenacity shortly before the molt as observed in Argiope aurantia presumably reduces the chance that a spider will be off the web when it begins to molt. Both mechanical difficulties and predation on the spider would be increased if the spider used neither retreat (Argiope uses only a web) nor web during ecdysis. Robinson and Robinson (1973) suggest that even a rudimentary web during the molt would reduce the risk of predation. Haskell and Moorhouse (1963) demonstrated a direct effect of ecdysone upon the peripheral neurons of an insect. But change of web-site tenacity is probably related to a central suppression of output from nerve ganglia for “dispersal” behavior rather than to direct effects on peripheral motor fibers, since spiders move their limbs and walk about the web even on the day of the molt.
Though larger Argiope aurantia change web site less often than smaller individuals, the distance between sightings of marked individuals, from one web site to the next, was greater the larger the spider. Thus, the average for 40 changes of web site of adult females was 5.1 m, while the average for small immatures was less than 1 m. Large subadult Araneus cornutus (Araneidae) on a fishing pier averaged 4.6 m in 30 changes of web site. But, smaller spiders are capable of travelling through the air (“ballooning”; Kaston, 1948; Enders, 1973), so it is possible that greater distances were travelled by smaller animals but that they simply could not be found at great distances due to the spiders’ small size (and the relatively large areas to be searched).
Thus, there are two significant results of this study: (1) change of web-site tenacity with molting, and (2) change of web-site tenacity with age or size. These effects are superimposed on a background of high web-site tenacity. At present, neither the internal physiological causes nor the ecological significance of these effects can be satisfactorily explained.
Acknowledgments.—This research was in part supported by NSF Grants GB-6246 to P. N. Witt and GB-27152 to W. F. Blair, being part of a thesis for
490
The American Midland Naturalist
94(2)
the Ph.D. degree in zoology at North Carolina State University. H. W. Levi identified the species studied. M. P. Clyde, A. C. Hulse, D. C. Lowrie, S. E. Riechert, J. S. Rovner and an anonymous reviewer made helpful comments on the style of writing.
Literature Cited
Blanke, R. 1972. Untersuchungen zur Oekophysiologie und Oekethologie von Cyrtophora citricola Forskal (Araneae, Araneidae) in Andalusien. Forma Functio, 5:125-206.
Breed, A. L., V. D. Levine, D. B. Peakall and P. N. Witt. 1964. The fate of the intact orb web of the spider Araneus diadematus Cl. Behaviour, 23:43-60.
Eberhard, W. G. 1971. The ecology of the web of Uloborus diver sus (Araneae: Uloboridae). Oecologia, 6:328-342.
Enders, F. 1972. Web site selection by Argiope aurantia Lucas and other orb weaving spiders (Araneidae). Ph.D. Thesis, North Carolina State Univ., Raleigh, N.C. 168 p.
1973. Selection of habitat by the spider Argiope aurantia Lucas (Araneidae). Am. Midi. Nat., 90:47-55.
-. 1974. Vertical stratification in orb web spiders and a consideration of
other methods of coexistence. Ecology, 55:317-328.
Haskell, P. T. and J. E. Moorhouse. 1963. A blood-borne factor influencing the activity of the central nervous system of the Desert Locust. Nature, 197:56-58.
Kaston, B. J. 1948. Spiders of Connecticut. Conn. State Geol. Nat. Hist. Surv. Bull., 70:1-874.
Klopfer, P. H. 1965. Behavioral aspects of habitat selection: a preliminary report on stereotypy in foliage preferences in birds. Wilson Bull., 77 : 376-381.
Krishnakumaran, A. and H. A. Sghneiderman. 1970. Control of molting in mandibulate and chelicerate arthropods by eedysones. Biol. Bull., 139: 520-528.
Levins, R. 1968. Evolution in changing environments: some theoretical explorations. Princeton Univ. Press, Princeton, N.J. 120 p.
Lowrie, D. C. 1963. Effects of grazing and intensive collecting on a population of the green lynx spiders. Ecology, 44:777-781.
Reed, C. F., P. N. Witt and M. B. Sgarboro. 1969. The orb web during the life of Argiope aurantia (Lucas). Dev. Psychobiol., 2:120-129.
Robinson, M. H. and B. Robinson. 1973. The ecology and behavior of the giant wood spider Nephila maculata (Fabricius) in New Guinea. Smithson. Contrib. Zool., 149:1-76.
Sale, P. F. 1969. Pertinent stimuli for habitat selection by the juvenile Manini, Acanthurus triostegus sandvicensis. Ecology, 50:616-623.
Snedecor, G. W. and W. G. Cochran. 1967. Statistical methods. Iowa State Univ. Press, Ames, Iowa. 593 p.
Turnbull, A. L. 1964. The search for prey by a web-building spider Achaearanea tepidariorum (C. L. Koch) (Araneae, Theridiidae). Can. EntomoL, 96:568-579.
Wecker, S. C. 1963. The role of early experience in habitat selection by the prairie deer mouse, Peromyscus maniculatus bairdi. Ecol. Monogr., 33: 307-325.
Witt, P. N., C. F. Reed and D. B. Peakall. 1968. A spider’s web. Springer Verlag, Berlin. 107 p.
Frank Enders,1 North Carolina Division of Mental Health Services, Research
Section, Raleigh 27611. Submitted 22 April 1974; accepted 16 December 1974.
1 Present address: Department of Zoology, University of Texas, Austin 78712.
Reprinted from
THE AMERICAN MIDLAND NATURALIST Vol. 94, No. 2, October, 1975, pp. 484-490
University of Notre Dame Press Notre Dame, Indiana


