Warning: Trying to access array offset on value of type null in /home/elisew5/public_html/drpeterwitt.com/wp-content/plugins/contextual-related-posts/includes/content.php on line 49
Images Collection
View this article in Search Friendly Plain Text
NOTE: This plain text article interpretation has been digitally created by OCR software to estimate the article text, to help both users and search engines find relevant article content. To read the actual article text, view or download the PDF above.
Fine Structure of the Thread Connections in the Orb Web of Araneus diadematus
BY ROBERT R. JACKSON-
Reprinted^op^PsYCHi^Vfi. $2, March-June 1971
pages 12-31
Reprinted from Psyche, Vol. 78, No. 1-2, 1971
FINE STRUCTURE OF THREAD CONNECTIONS IN THE ORB WEB OF ARANEUS DIADEMATUS1
By Robert R. Jackson2 Division of Research North Carolina Department of Mental Health Raleigh, North Carolina
INTRODUCTION
Web-building spiders are valuable subjects for the study of behavior since the spider provides a record of much of its behavior through its web. Descriptions of the gross structure of the web and the behavior of the spider during its construction can be found in Savory (1952), Jacobi-Kleemann (1953), and Witt, Reed, and Peakall (1968). However, the gross structure is only part of the spider’s creation. The threads from which the web is constructed are only several microns thick. Consequently, the unaided human eye can determine basically only the position of the threads.
Another part of the spider’s creation is visible only with magnification. This is the fine structure of the threads where they connect to one another. Rapidly produced and possessing remarkable strength (DeWilde, 1943), thread connections are one of the more interesting accomplishments of the spider. However, little attention has been given to the fine structure of the web. Lehmensick and Kullmann (1957) and Friedrich and Langer (1969) examined spider silk by electron microscopy, but did not look at points of connection between threads. McCook (1889) and Nielsen (1931) have drawings of attachment disks for drag-lines made under low magnification. De-Wilde (1943) has a photomicrograph of a radius to frame connection, and Comstock (1948) has a drawing from a photomicrograph of a spiral to radius connection. Robbins (Savory, 1952) has a photomicrograph of a drag-line attachment disk and one of sticky spiral to radius connections, both at low magnification. Also, there is a photomicrograph of a spiral to radius connection from the hori-
This work was carried out in the laboratories of the North Carolina Department of Mental Health and was supported in part by a grant from Hoffman-LaRoche, Inc., Nutley, New Jersey, and by Grant Number GB-6246X1 from the National Science Foundation to Peter N. Witt. The assistance of Dr. Peter N. Witt during all stages and Dr. Charles Walcott during the preparation of the manuscript of this work is gratefully acknowledged.
2Mailing address: Robert R. Jackson, Department of Zoology, University of California, Berkeley, California 94720.
12
1971]
Jackson — Web of Araneus
13
zontal web of Cyrtophora citricola in Kullmann’s (i957) study of this peculiar spider. However, we have been able to find no detailed descriptions of thread connection fine structure.
In the study reported in this paper, we investigated the fine structure of most of the types of thread connections which are found in the web of Araneus diadematus Clerck. Within the web there may be between 1000 and 1500 locations at which one thread is fastened to another thread (Fig. 1). Mooring threads fasten the web to some non-thread structure and may be continuous with some of the frame threads (Witt, et aL, 1968). There are frame thread to frame thread connections, forming Y’s with the stem of each Y being a mooring thread. Each radial thread is fastened to a frame thread. Occasionally, a radius will be in the form of a Y-structure at its peripheral end; the stem of the Y connects with the hub; and each arm, or secondary frame thread (Peters, 1939), fastens to either a frame thread or a radial thread. However, the majority of the connections within the web are the viscid or sticky spiral to radius connections in the trapping zone. Most of these are at points at which the sticky spiral thread extends from both sides of the radius. Occasionally, especially in the lower portion of the web, the sticky spiral thread meets the radius but does not continue on the other side. In the strengthening zone (Savory, 1952) or notched zone (McCook, 1889), non-sticky spiral threads are fastened to radial threads. The strengthening zone surrounds the apparently disorderly network of threads in the hub (McCook, 1889). Non-sticky spiral thread to radial thread connections are also present in the provisional spiral, a structure which is removed as the sticky spiral is added. Each of these types of connections will be described except for those at which a sticky spiral meets the radius but does not continue on the other side.
METHODS
All thread connections surveyed in this study came from webs of laboratory reared adult female cross spiders (Araneus diadematus Clerck). Each spider was kept in a 51 X 51 X 9 cm aluminum frame with removable glass doors and was provided with one house fly twice per week, water daily, and controlled conditions of light and temperature (Witt, et al., 1968). The age (3 to 4 months since emergence from the egg cocoon), weight (88 mg to 134 mg), and time between food consumption and web construction (1 to 8 days, with the majority at 1 day) were relatively constant for spiders used in this study; and within these ranges for these factors, no cor-
14
Psyche
[March-June




19?1] Jackson — IVeb of Araneus 15
relation was apparent between these factors and structural features of the connections.
The day before the connections in a spider’s web were to be looked at, the glass doors were removed, and the old web was destroyed. The new web, built by the spider early the next morning, was treated as in Witt et al. (1968) except that the threads were not coated in any way. Afterwards several thread connections from the web were examined.
In the web, threads are stretched tight into straight lines. When placed on a glass slide, threads with sticky globules often remained straight; but the globules were usually destroyed. Threads without globules curled and twisted when placed on a slide. To obtain a less distorted connection to study under a microscope, Permount mounting medium (Fisher Scientific Company) was streaked into a circle on a glass microscope slide, forming a basin. The slide was then placed against the web so that the threads surrounding the connection would stick to the Permount. Thus, the connection was suspended above the slide by the ridges of the circle of Permount. The threads were then burned loose from the outer edges of the circle by a surgical cauter, and the slide was taken away. The slide was always made on the same day that the web was built.
Within a week of preparation of the slide, the connection was viewed under a Leitz Labolux D microscope; and negatives were made on Panatomic X sheet film using a Brinkmann Mark V camera with various objective lenses on the microscope. That structural
Fig. 1. The vertical orb web of Araneus diadematus with labeled structures. The scale, consisting of a weight suspended by cotton threads 20 mm apart, indicates size and vertical direction. Abbreviations: SS, loop of sticky spiral; SS-R, sticky spiral to radius connection; NS-R, non-sticky spiral to radius connection; R-F, radius to frame connection; FY, frame Y-structure; RY, radial Y-structure.
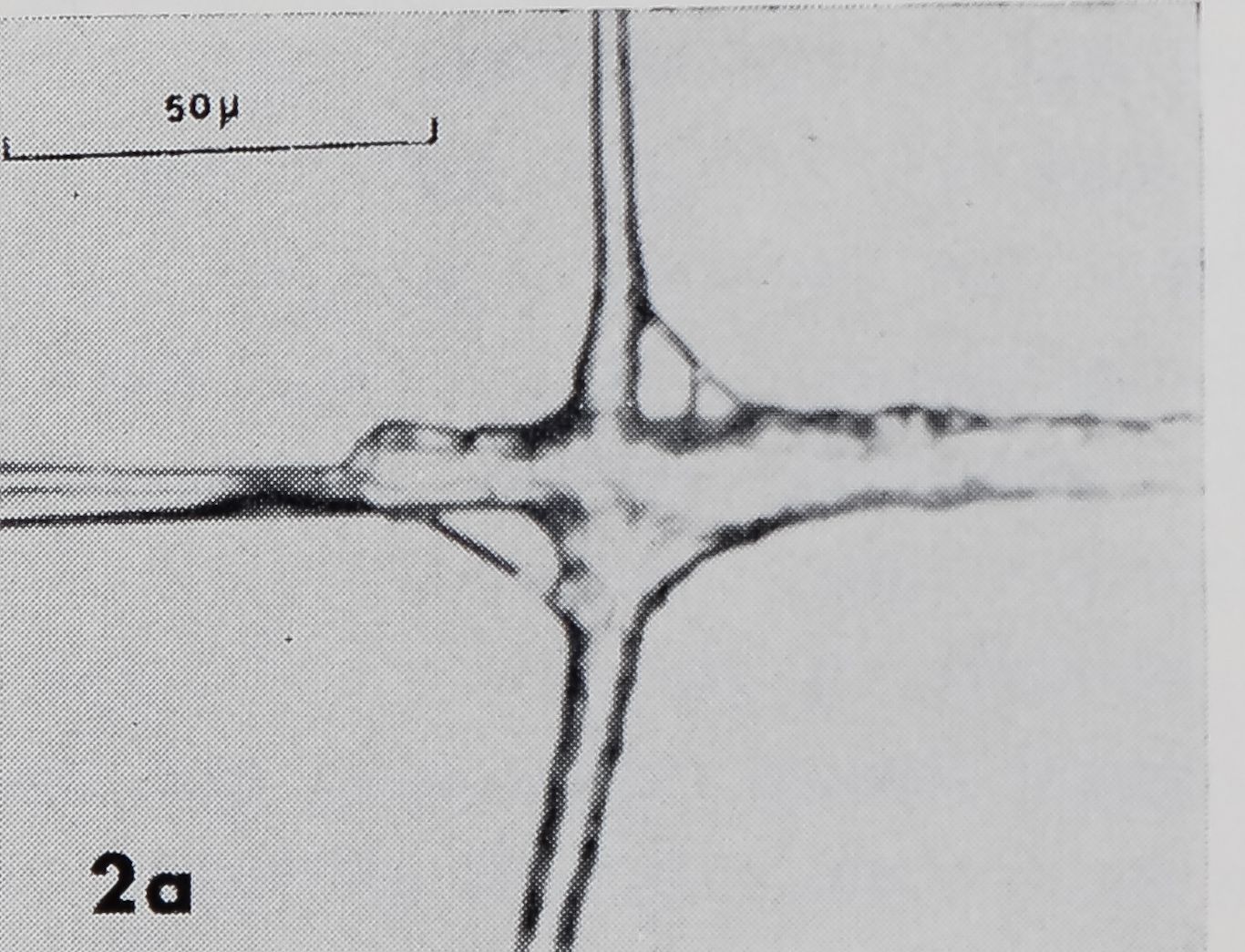
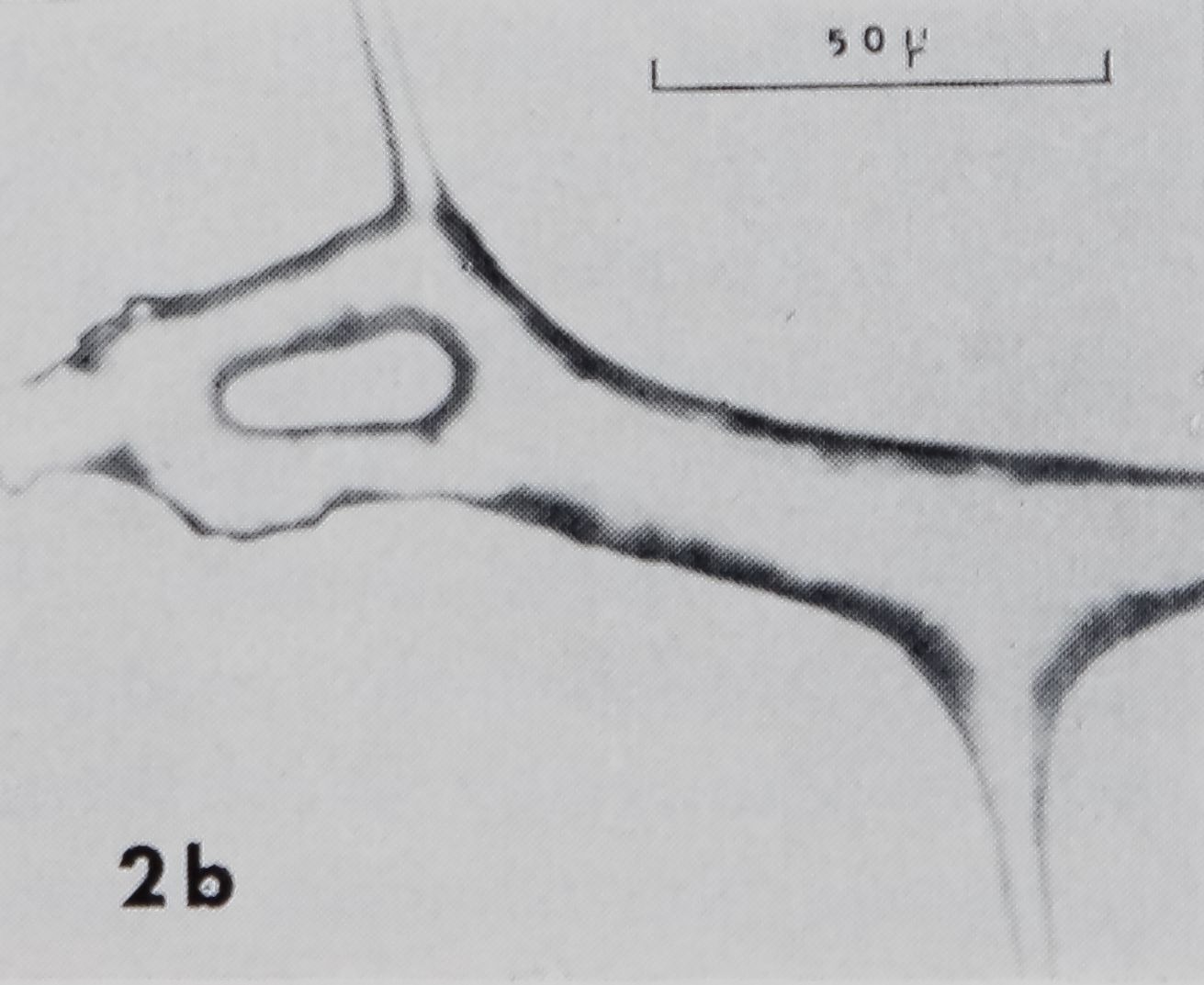
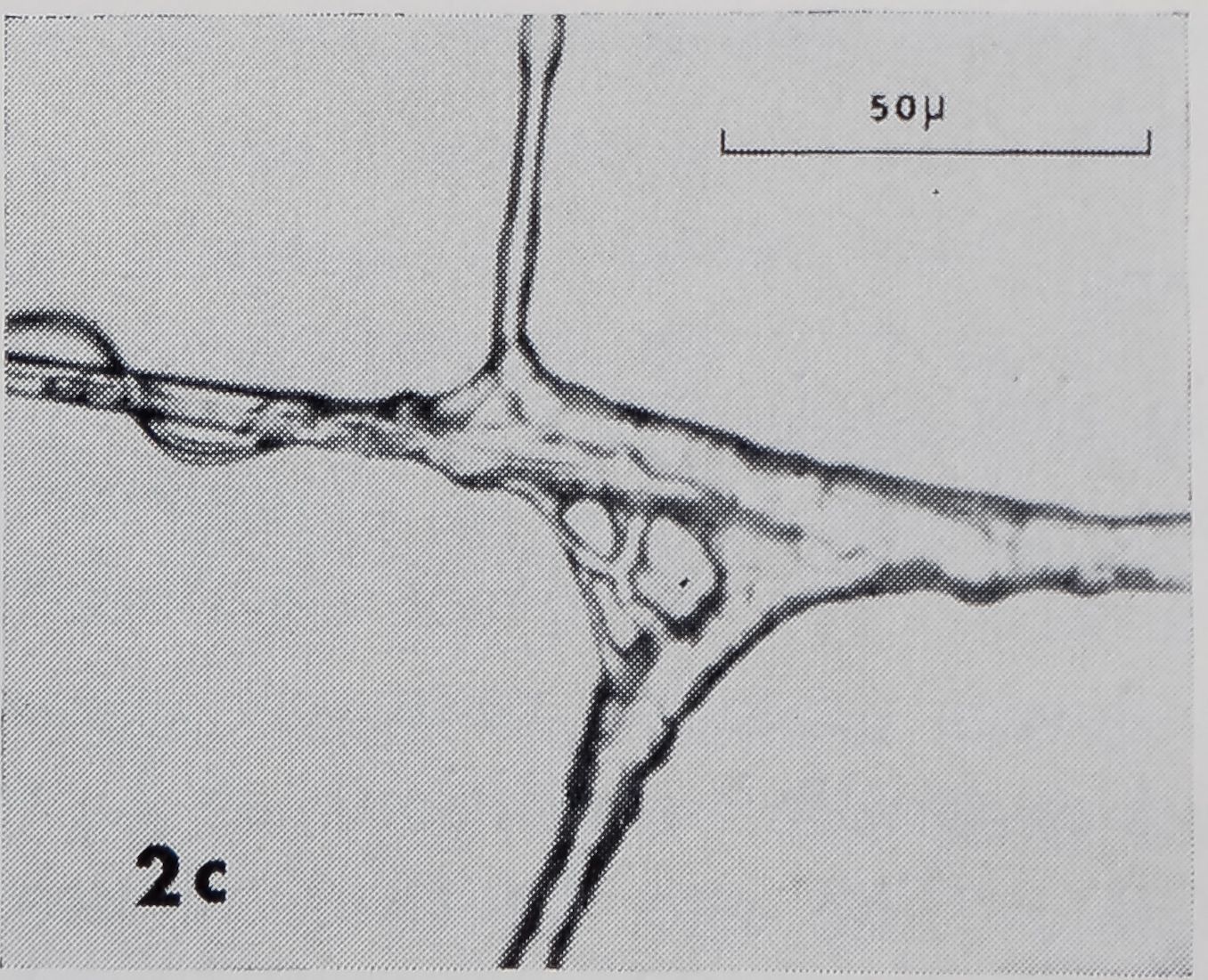
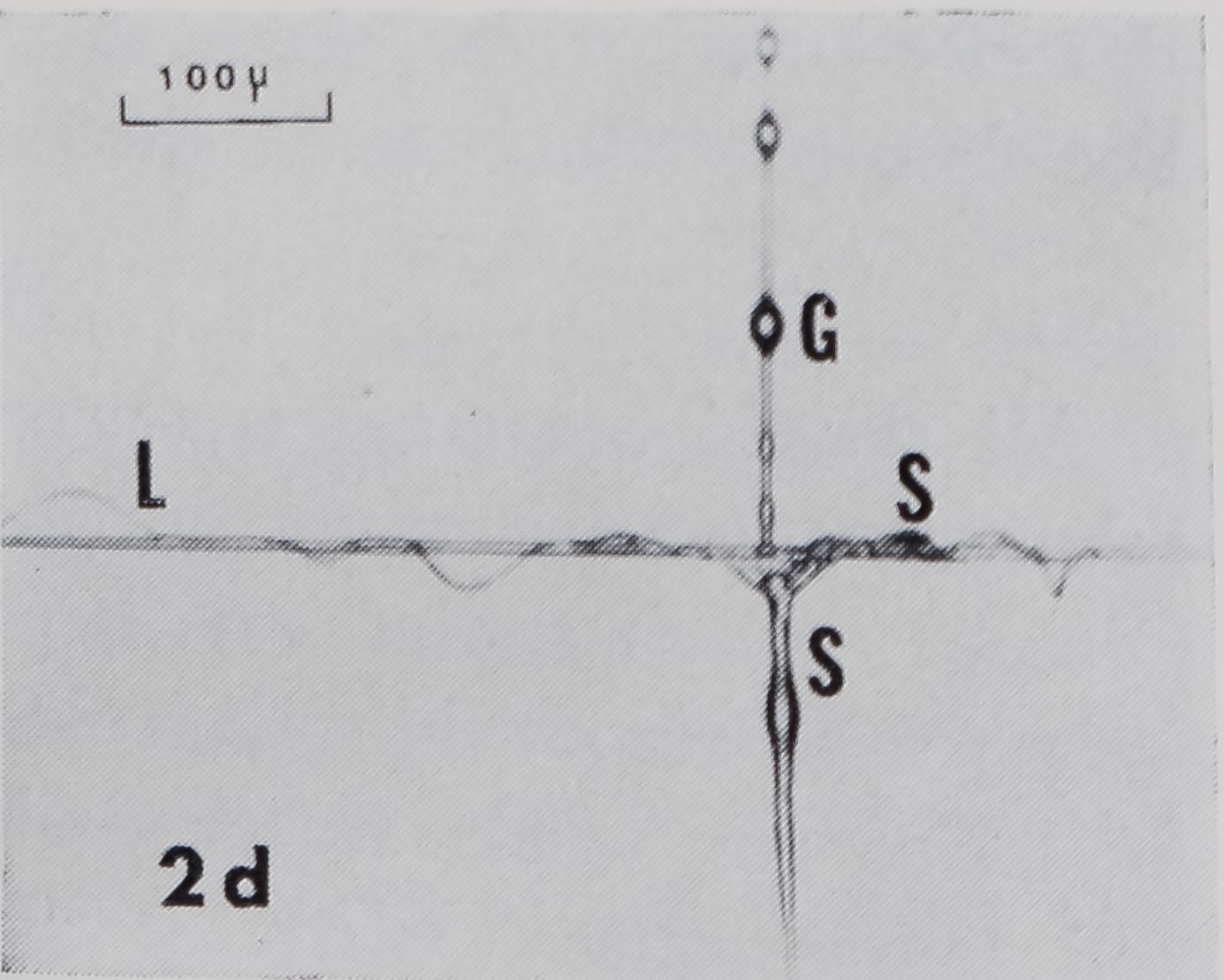
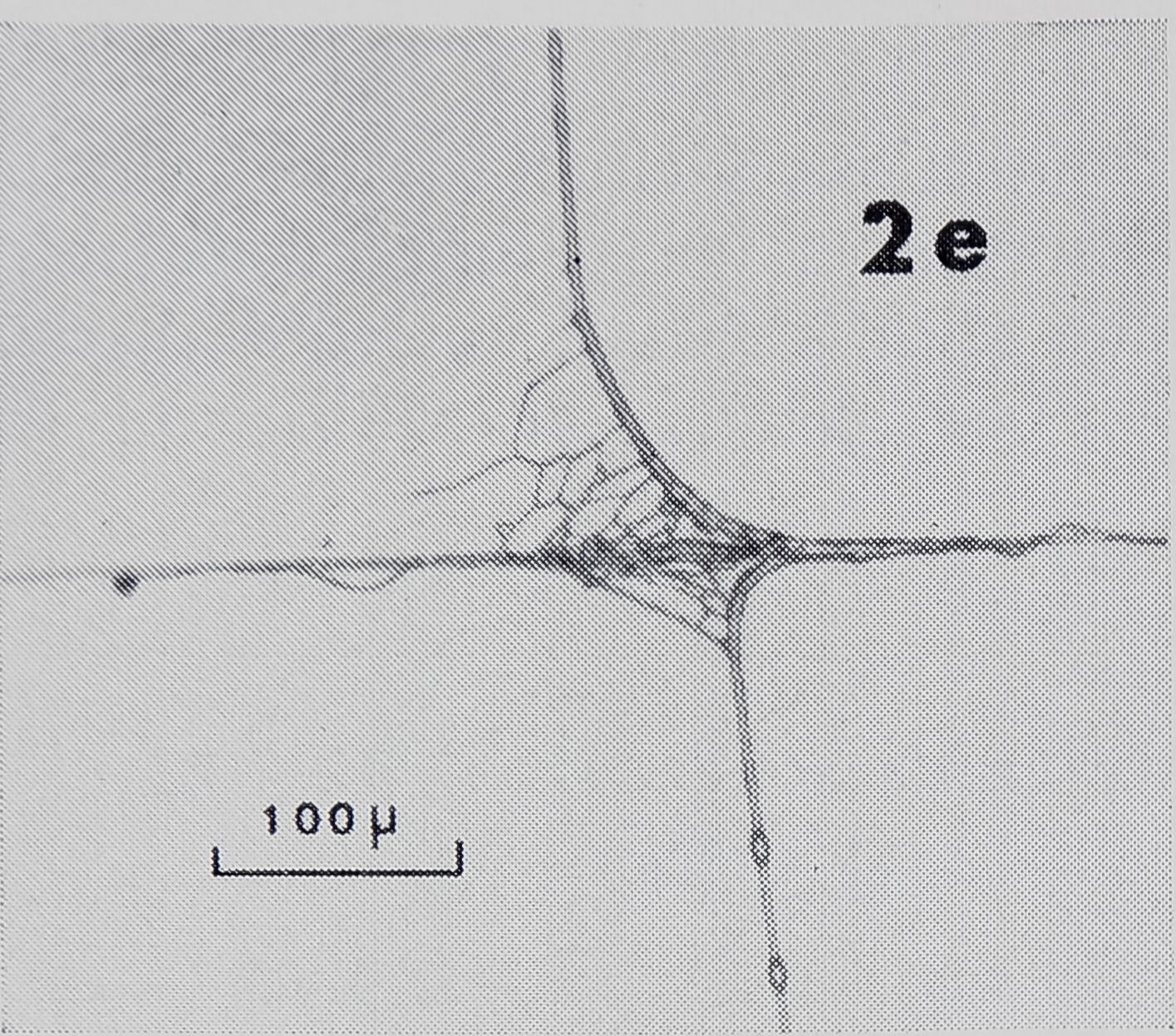
Fig. 2a-d. Examples of sticky spiral (SS) to radius connections. In these photomicrographs, SS’s run vertical ; radii, horizontal. Radius is fastened to frame to right of page. Up and down on page corresponds to up and down, with respect to gravity, in web. a. Zero junction: points where SS joins radius on one side is directly across from where it joins at other side. Note sleeves on radius at both frame and hub side of SS. b. Smooth non-zero junction: there is space, smooth in appearance, along radius between points at which SS joins each side of radius. Idiosyncrasies such as the doughnut-shaped structure in this SS-R made generalization about structure difficult, c. Non-zero junction with rough appearance, d. Note sleeves (S) on radius at frame side of SS and on SS, loose strands (L) on radius, and globules (G) on SS.
lb
Psyche
[March-J univariations of the connection were not due to the amount of time which elapsed between slide preparation and exposure of the negative was shown by comparing photomicrographs with the slides after more time had elapsed. After several weeks to a year, essentially no changes in structure were discovered except for frequent disappearance and fusion of globules on the sticky spiral.
Various measurements and counts were made from the negatives and prints. Since I [x was close to the limit of resolution on the photomicrograph, measurements in this range should be considered approximations. The thickness of a thread associated with a connection was usually measured at a distance of 150 from where the thread joined the other thread. In most cases, threads were fairly uniform in thickness at this distance. Closer to the other thread, thickness had a greater tendency to vary, and measurements were complicated by features of the connection. When the thickness of a thread at 150 [x was obviously not typical for that thread, as for example, when a sticky spiral had a globule on it at this point, the thickness was measured somewhat further from the connection. When strands were counted at a connection, “separate strands” were defined as strands with a visible space between them in the photomicrograph. However, sometimes an estimation of the number was used because some strands were hidden by others, and some were out of focus.
The reader may refer when necessary to the following list, in alphabetical order, of abbreviations which will be used in this paper; FY, frame Y-structure; NS, loop of non-sticky spiral from the strengthening zone; NS-R, non-sticky spiral to radius connection from the strengthening zone; PS, loop of provisional spiral; PS-R, provisional spiral to radius connection; R-F, radius to frame connection; RY, radial Y-structure; SS, loop of sticky spiral; SS-R, sticky spiral to radius connection.
RESULTS
The microscope reveals a wealth of structural detail at thread connections from which many component features can be abstracted, and the complexity and variability of which make description difficult. Some features of the structures will be pointed out in the following sections; and sample photomicrographs will be presented. Various descriptive statistics (percentages, means, coefficients of variation, and ranges) will be used to provide an impression of the prevalence of certain features and the central tendency and variability of mens-
1971]
Jackson—Web of Araneus
17
Table 1
Frequency of some structural features of sticky spiral to radius connections (SS-R’s). For each feature, 49 SS-R’s from IS webs built by 8 different spiders were observed. Note that each SS-R had a sleeve on the radius at the frame side of the junction and that SS’S never had loose strands.
| Feature | Percent of SS-R’s having the feature |
| Zero Junction | 22.4 |
| Sleeve on Radius at Frame Side of Junction | 100 |
| Sleeve on Radius at Hub Side of Junction | 46.9 |
| Sleeve on SS | 55-i |
| SS and Radius Both Single Stranded at Junction | 36.7 |
| Loose Strands Along Radius | 35-5 |
| Loose Strands Along SS | O |
urements made on various features. We will draw attention to some of the questions suggested by these observations, and possible directions for further research will be indicated with hypotheses concerning function and origin of the structures.
Sticky Spiral to Radius Connections:
In reading the following section, the readers may refer to Table 1 and Table 2 for data pertaining to SS-R’s.
We will define a “junction” as the space along a thread at which another thread is fastened to it. By “connection” we will mean
iS
Psyche
[Marcb’Juoethe junction plus the threads in the immediate vicinity of the junction.
The point at which the SS joined one side of the radius was either directly across from where it joined the other side (zero-junction) (Fig. 2a) ; or there were varying amounts of space (the junction) along the radius between these points (non-zero junction) (Fig. 2b), causing the SS to have a disconnected appearance. In contrast to the SS’s, radii always appeared as continuous, nearly straight lines at SS-R’% Junctions were smooth (Fig.’-à^por rough (Fig. 2c), with many gradations j§é£^en, The smoother junctions were more prevalent.
It would be interestmgMj know inpp|pt if any, the length and roughness of the art n3^^;^:‘tlp^Hç’ttoning of the
web. -the con
nection’ or the’
Th«f”ffliuS ^ ai^-R’sWd, rar%equently,
the radius at the MyJMpI»’ and
jft- fife junction than the same ‘^^Hpwb^f^w’ay from the junction. These areas will be ig-
2qy.,f; JfWany gi\lep’junction, tHHHpl&!f^ the radi*^Bthe frame side #Ms at^Alfel as (± the tfc hub
side; and u-sullMS’%is fcngef than the one on 8b^S$ ‘
tjfedlds mm strengthened lj$ ftp$iHpj&nd tbfe^flpâ^nce and length of’ tJIteyeSj ‘ are- related to the magnitudë«p the tensions th^fp;must withstand in the web. 3V|fc^suremgntfe|f thes|.^^^fc;, would be some SS Mn|^the radius as a
single strand at both ^[Fî^lb and Other
•ffi-R’s^c’onsis’ted ^^fe^feiinbers of strands i§!Fig. jtedMWt <2e), often int»çpn®^&| gnd variable nKbickdess. Perhaps, the pÉSrajjpe of many a junction functions in the dispersal of stresses
or ter increase elasticity.
If: the presence junctions, lipftijfg on the
radii at the hub side of junctions, sleeves on SS’s, and many strands at the çp^iect^mthe web^Esome wam^^^^Sfe^iome SS-R’s
have these characteristics while other|M^.kngt.? ,
Many SS-R’s had jdiosyn.Qra^^<|the description ojr. which will not be attempted here. The doughnut ‘ shape! Structure in Fig 2b is an example. It does hot ‘seem likely that these structures have a specific function since any one of them occurred at only a single SS-R. They are probably artifacts of the forces acting on the connection during its formation. Thread connections, especially SS-R’s, are produced at a very rapid rate. The spider produces over a thousand
Table 2
Descriptive statistics for some structural features of sticky spiral to radius connections (SS-R’s). SS-R’s were rather variable with respect to these features, as indicated by the coefficients of variation. Note the differences in sleeve length on the radius at the different sides of the junction and the differences in thread thickness for the radius and SS. Thread thickness was measured at both sides of the junction.
| Coefficient | Number of | Number of | Number of | |||
| Feature | Mean | of Variation | Range | Measurements | Webs Examined | Spiders Used |
| Length of Junction | 27.8 p | 98 | 0 At-100 p | 49 | 15 | 8 |
| Length of Sleeve on Radius at Frame Side of Junction | 106.6 n | 39 | 37 p-200 p | 48 | 15 | 8 |
| Length of Sleeve on Radius at Hub Side of Junction | 57.2 P | 54 | 25 p-100 P | 23 | 9 | 8 |
| Length of Sleeve on SS | 43.6 p | 59 | 7 p-125 p | 37 | 13 | 6 |
| Number of Strands at Junction | 10.0 | 100 | 4-42 | 49 | 15 | 8 |
| Distance Between Junction and Nearest Globule | 49.3 P | 52 | 6 a*-112 p | 37 | 11 | 6 |
| Number of Globules Within 300P of Both Sides of Junction | 6.5 | 54 | 0-12 | 49 | 15 | 8 |
| Thickness of Radius | 5.8 p | 16 | 3 P-9 p | 96 | 15 | 8 |
| Thickness of SS | 3.7 ii | 25 | 3 P-5 p | 98 | 15 | 8 |
20
Psyche
[March-JuneSS-R’s in approximately ten minutes. Taking this into consideration, it is not surprising that there is a great deal of variation in connection structure and that SS-R’s «J|||P^feefaiiés. ;
Globules (Fig. 2^ftpresumably containing the viscid material, were hut none were seen
0$ radii.|||^Bp^^^M§^SlH^M^iEnt of space between the junction anObservations of p||^lÉH9^ions other than . impression that the space
within a given length of thread) constant. In contrast, the number oSfflobules
wipln; of ft W-R was quite variable.
Stretching the Sgis the nmhable material; inldT
through being lliililonfr]nijrnv
It gBragljlPed thaw^^^wâiïlB^t^lÉl^ thei
radii and glands producing
tfye viscid material ( Peakall,3^^^HMd)0|çÉp^^^^^^^HHg|ny not simply equivalent to a viscid spiral thread minus the viscid ntaterial’i radial threads we observed were
thicker than SS’s, although both li§9- the same minimum thickness.
; ^Mai^iéiw ilde ( 1943 ) found
more extensible than radi^^^m^^SM^-^d^ÔSrved’ loose strands
Zone and Provisional Spiral to
Radius Connections t’Ç
Table 3 and ffiaflHj4 NS-R’s^,. ^he
motion pattern of the N S.-R’s
and the other* The fourth
pair of legs are and fMchingj,«the radius as a
NS or is fastened to a radius. However, .âfllheuspinnerets touch the radins in generating ii j lKR, one., of the fourth legs stretches the SS. In comparison t<ÿ;‘:M|j-R’s and PS-R’s the construction of SS-R’s apears’UbW and deliberate..jÇfitferorv. 1952®® There are also differences in the fine structure of these connnections.
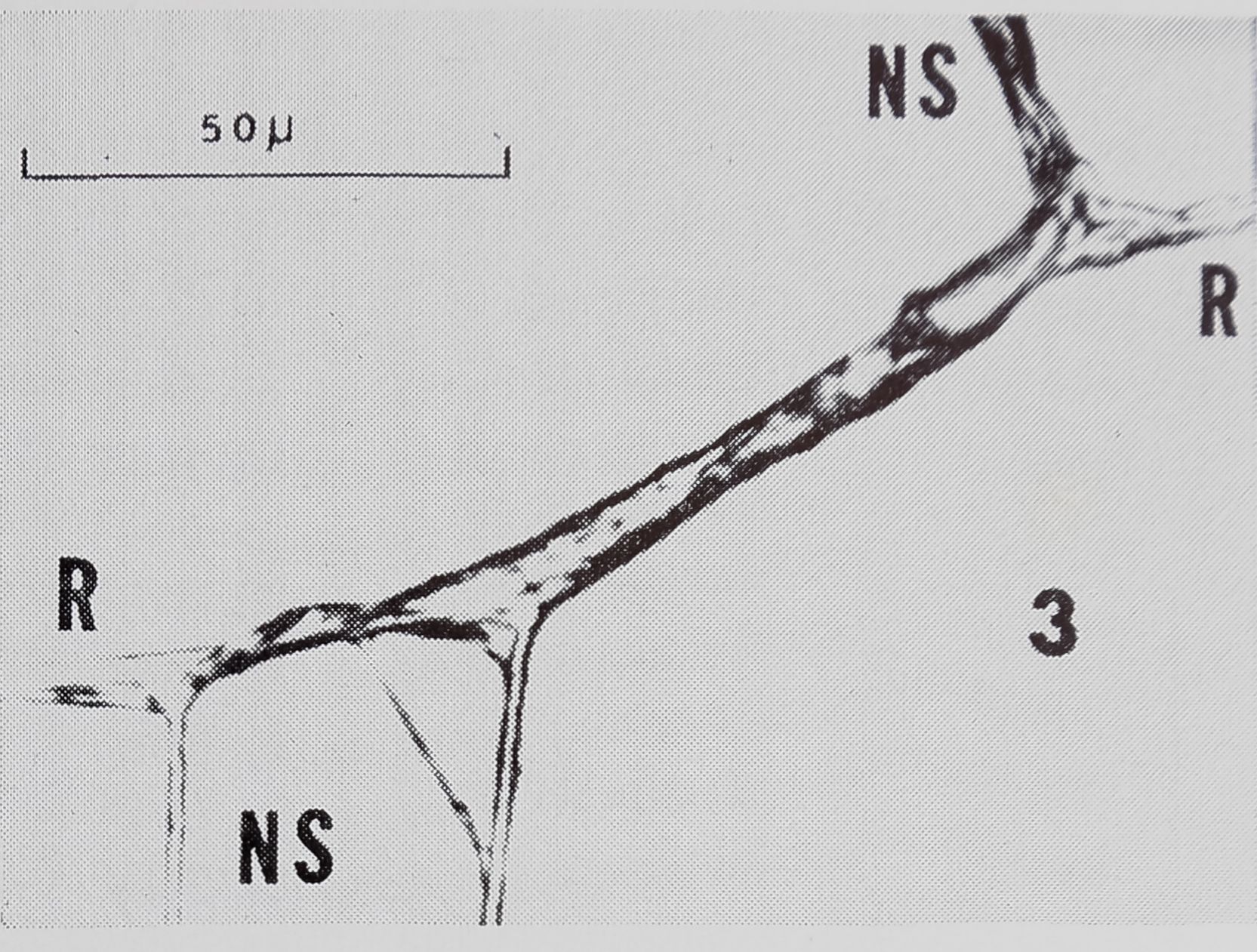
Junctions of NS-R’s- (Fig. 3) were usually much longer than those of SS-R’s. There were no zero junctions. In contrast to SS-R’s, neither radial nor spiral threads ever appeared as continuous lines at NS-R’s. This created the notched appearance of the strengthening
1971]
Jackson — Web of Araneus
21
Table 3
Frequency of some structural features of connections from the strengthening zone (NS-R’s). For each feature 18 NS-R’s from 2 webs built by different spiders were observed. Comparing this table with Table 1, note the absence of zero junctions, the decreased frequency of sleeves on radii, and the presence of loose strands along the NS.
| Percent of NS-R’s having | |
| Feature | the Feature |
| Zero Junction | 0 |
| Sleeve on Radius | 5.5 |
| Sleeve on NS | 22.2 |
| NS and Radius Both Single | |
| Stranded at Junction | 5-5 |
| Loose Strands Along Radius | II.I |
| Loose Strands Along NS | II.I |
zone. Also, unlike SS-R’s, the junctions of NS-R’s always had a rough appearance.
NS-R’s had sleeves on radial or spiral threads less often than SS-R’s. Unlike some SS-R’s, there was a sleeve on no more than one strand at any one NS-R.
As at SS-R’s, a spiral thread at a NS-R often joined the radius as several strands. However, some SS-R’s had many more strands than were found at NS-R’s.
Only two PS-R’s (Fig. 4a)were looked at, and they resembled NS-R’s in length and roughness of the junction, in configuration of threads, and in having sleeves and a number of strands.
Is the manner in which the structure of NS-R’s and PS-R’s differs from the structure of SS-R’s related to differences in movement pat-
22
Psyche
[March-JuneFig. 2e. Sticky spiral to radius connections. Many strands connecting SS to radius.
Fig. 3. Typical non-sticky spiral to radius connection from strengthening zone. Sleeves can be seen on spiral (NS) at one side of radius (R) ; NS at other side consists of 3 strands. There is rough appearing space between points at which spiral joins each side of radius, and radius is out of line.
Fig. 4. Provisional spiral (PS) to radius (R) connection, a. Notice resemblance to NS-R’s (Fig. 3). b. Globules can be seen on one of the strands.




1971]
Jackson — Webof Araneus
2 3
terns during web construction? The spinnerets are capable of “to and fro” and complex rotary movements (Wilson, 1969). It would be interesting to observe these movements during the construction of each type of thread connection. For example, there may be differences in construction procedures on this level which can be related to the manner in which the fine structure of SS-R’s differs from that of NS-R’s and PS-R’s. Also such observations might provide information about the origin of some of the structures at thread connections.
The production of radii, NS’s, and PS’s, is attributed to the ampullate glands. The aggregate glands, to which production of the viscid globules is atributed, are not believed to be involved (Peakall, 1969). Some spiral and radial threads at NS-R’s and PS-R’s resembled radii at SS-R’s in having loose strands and no globules. However, one PS-R had tiny globules, smaller than those on SS’s on a thin (0.5 j u.— 1 jx) strand (Fig. 4b). How were these produced, and what glands were involved?
Table 4
Descriptive statistics for some structural features of connections from the strengthening zone (NS-R’s). For each feature 2 webs built by different spiders were observed. NS-R’s were rather variable with respect to these features, as indicated by the coefficients of variation. Comparing this table with Table 2, note the increased length of the junction and the decreased] thickn’ess of the radius. Thread thickness was measured at both sides of the junction.
| Feature | Mean | Coefficient
of Variation |
Range | No. of Measurements |
| Length of Junction | 137.4 fJ | 29 | 75 11-225 /i | 18 |
| Number of Strands at Junction | 8.9 | 49 | 4-22 | 18 |
| Thickness of Radius | 3.9 p | 41 | 1 /U-7 ju | 35 |
| Thickness of NS | 3.1 jW | 29 | 2 /u-5 1* | 32 |
24-
Psyche
[March-JuneTable 5
Descriptive statistics for some structural features of radius to frame connections (R-F’s). For each feature 5 webs built by 4 different spiders were observed. R-F’s were rather variable with respect to these features, as indicated by the coefficients of variation. Note that frame threads at R-F’s were thicker than threads at SS-R’s and NS-R’s (see Table 2 and Table 4).
| Coefficient | No. of | |||
| of | Measure | |||
| Feature | Mean | Variation | Range | ments |
| Length of Sleeve | 158.7/c | 38 | 100 /J-250 1* | 8 |
| Number of Strands in Radius | 34.1 | 24 | 19-45 | 8 |
| Thickness of Frame | 8.4/4 | 27 | 5 M-12/t | 13 |
It is interesting that a spiral to radius connection of another species, Cyrtophora citricola (Kullman, 1957) greatly resembled NS-R’s of A. diadematus. It had a similar configuration of threads, a non-zero junction, the spiral thread joining the radius as several strands, and a loose strand on the spiral thread.
The Hub:
When the spider completes the sticky spiral, she removes the threads at the hub and adds new ones. Here she sits when the web is completed and from time to time fastens more threads.
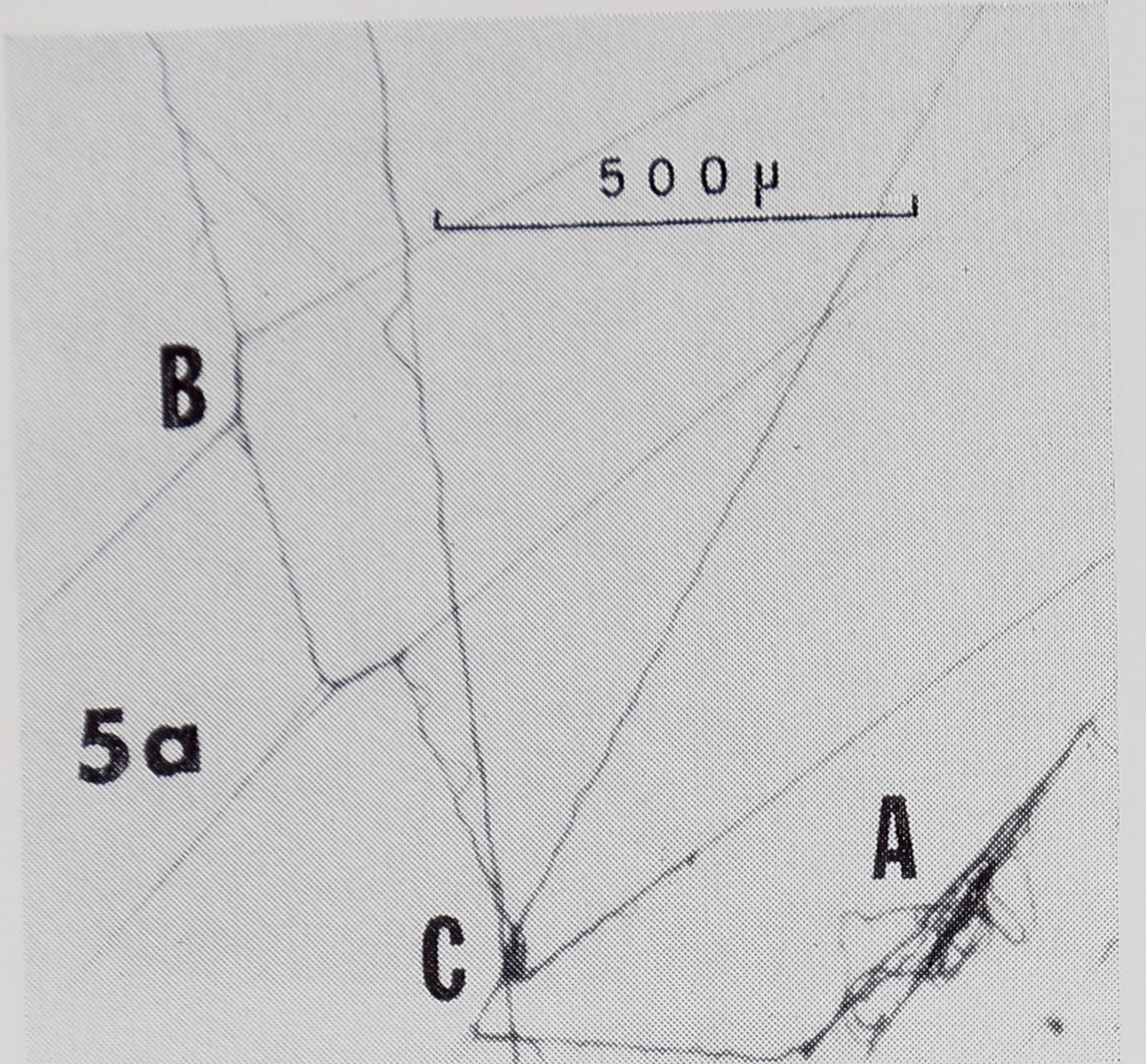
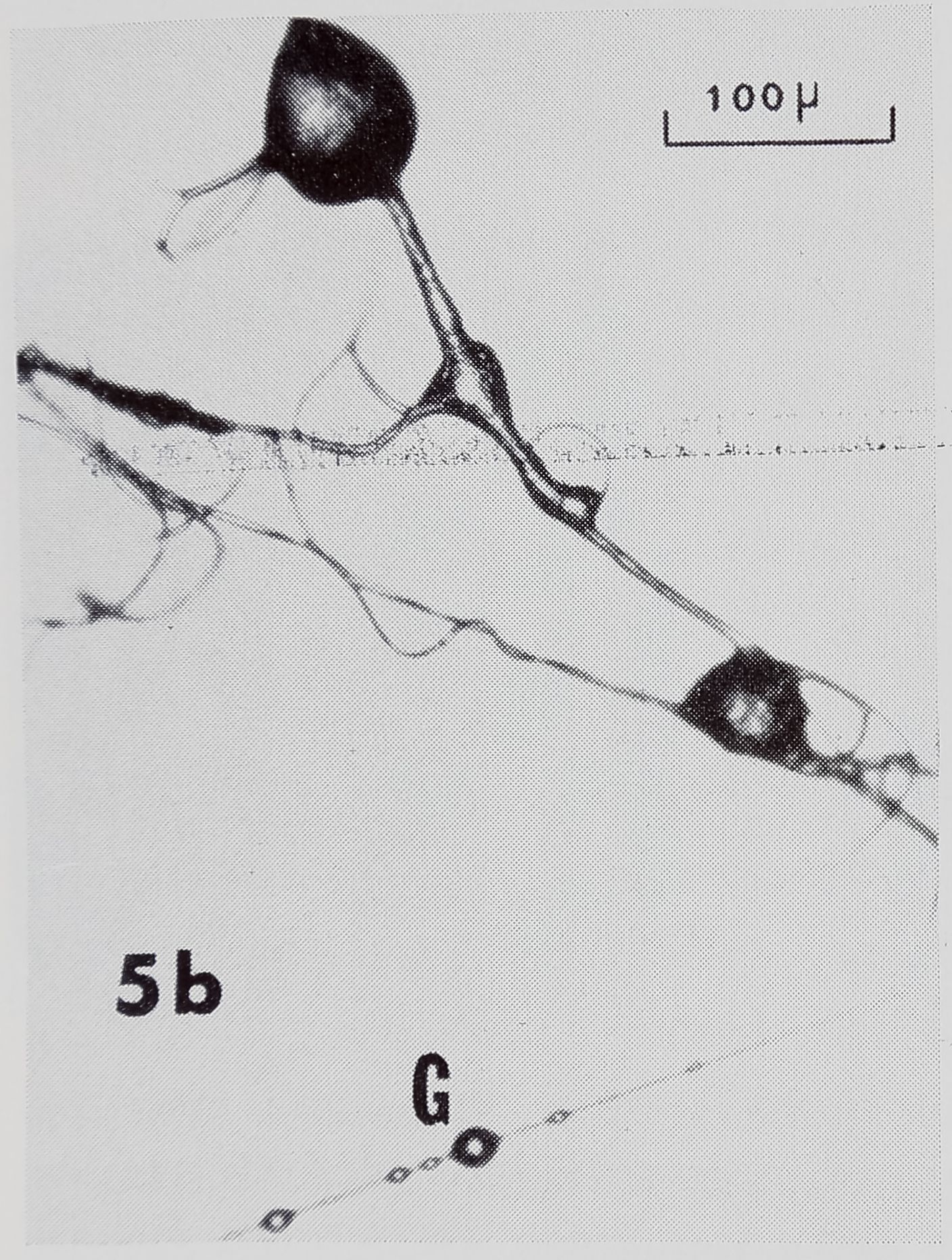
At the only hub we investigated, there were thread connections resembling NS-R’s (Fig. 5a). Others were compound, with threads radiating from the junction in more than 4 directions, probably the result of two or more connections being made at the same place at different times. The shapes of some junctions from the hub (Fig. 5b) were quite unlike the shapes of those from any other observed type of connection. There were many loose threads and strands of thread (Fig. 5a), perhaps the effect of changing tensions as the spider moved about the hub. The discovery of globules on threads
19?l] Jackson — Web of Araneus 25
at the hub (Fig. 5b) resembling those on SS’s, raises the question of which glands are in operation as the hub is formed.
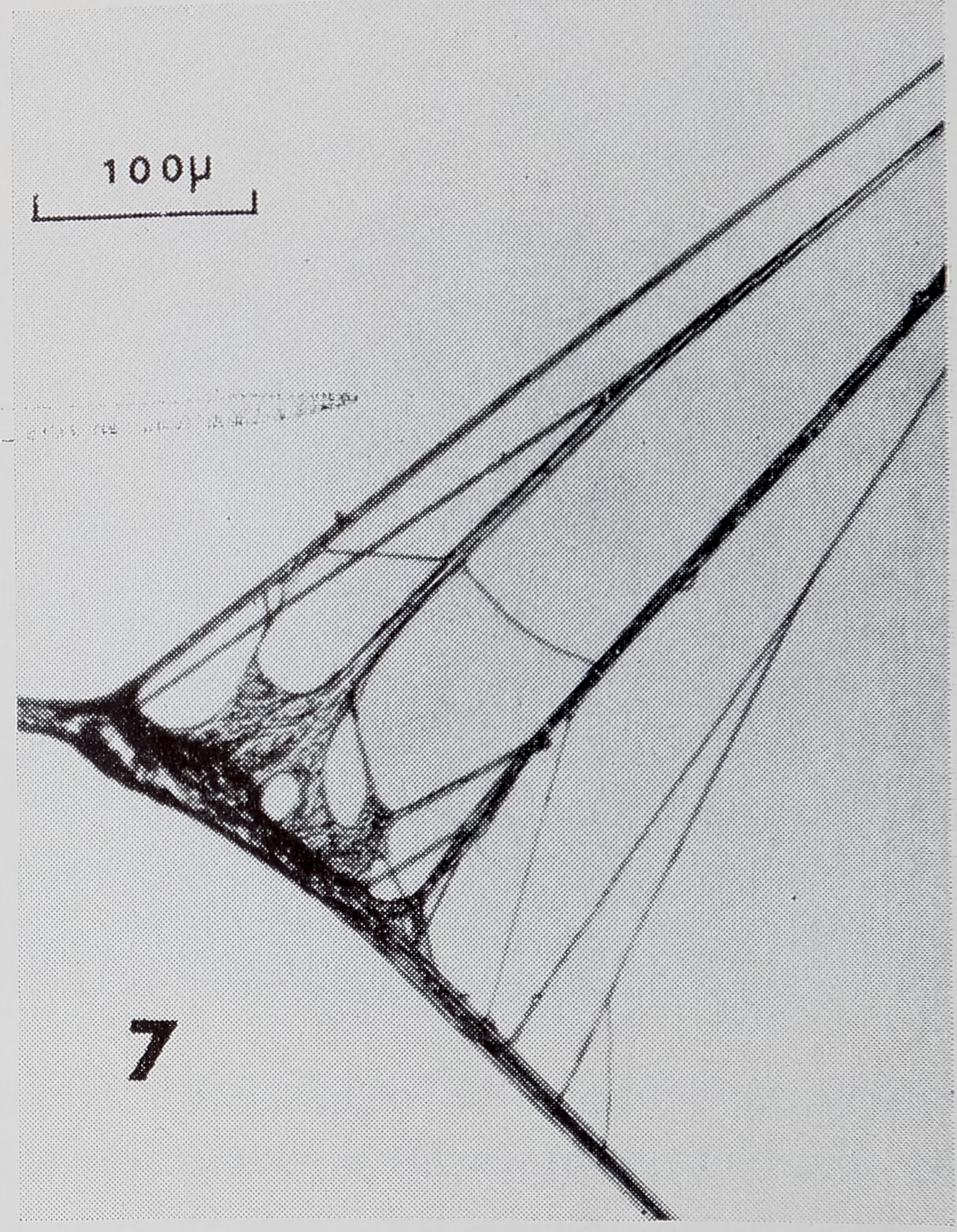
Radius to Frame Connections and Y-Structures:
In reading the following section the reader may refer to Table 5 (R-F’s) and Table In general appearance, R-F’s and
Y-structures resembled each other more than they resembled other type^JH. oççmbc^ons in web. However, many the structures we ¥fl|f• .A^^rtmentioneliW’Mfe^.«juisiyrt of otheF types of thread conneef^pSeii.. also? found at these connections.
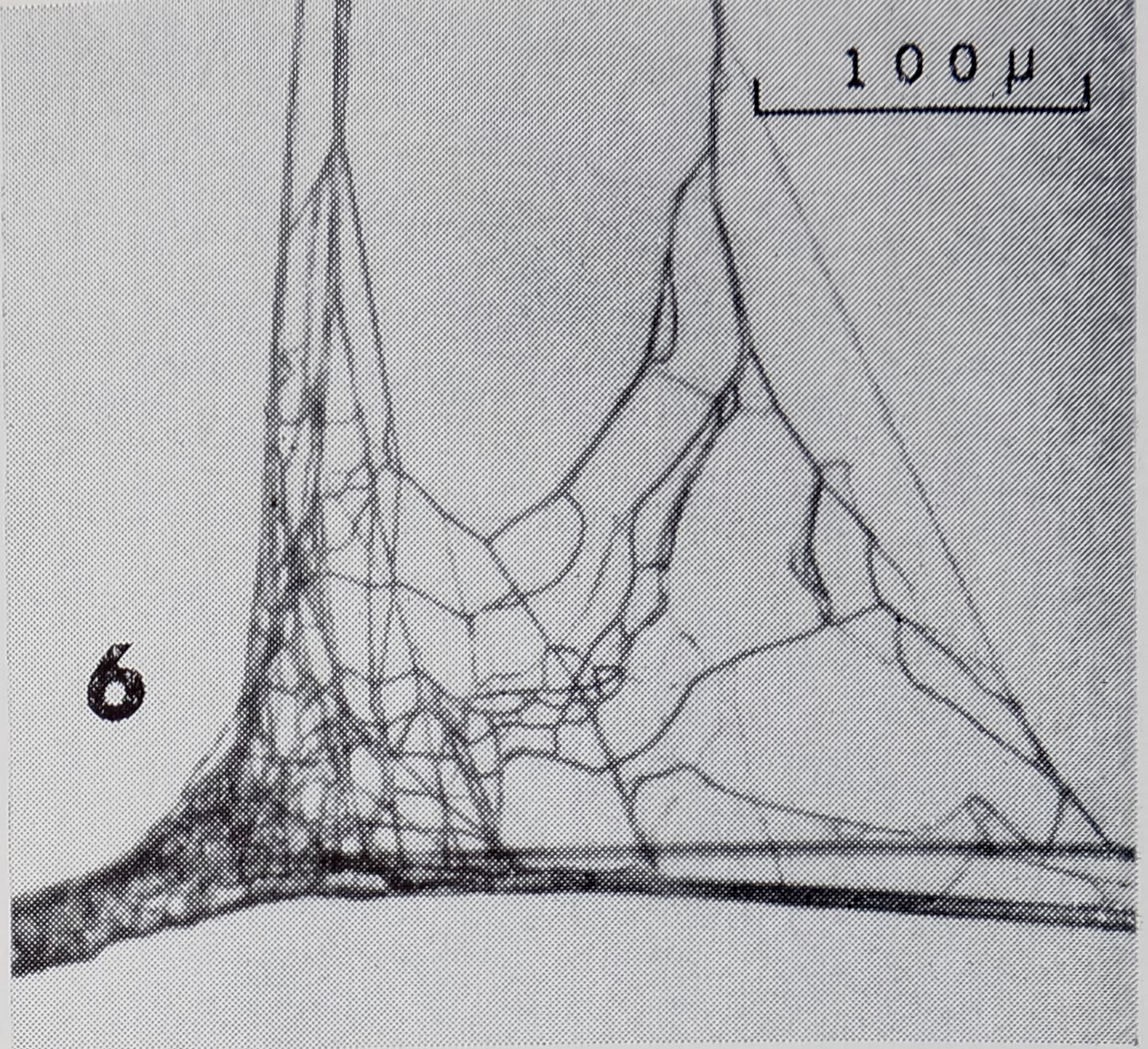
Wè examined 8 R>IBB^ RY. Each R-F (Fig. 6)
and FY| had one slceteèt Those at R-F’s were em the frame;
Item of the Y. The RY ,*^<Fig. 8) had 2 sleeves. One an arm. Sleeves at
R-Fs, FY’s and the RY lorï;g%or longer than those at
#|Im
R-F^fHB|the RY ^§t|SV<lt Each R-F
CQs^^^BHm|^WMp^Rtrand(^-t^-adÉà.’.MpP^^ to % single stranded ïo^^m(0′ whl^l. the frame was spli£ into 2 strands al one side of FY consisted of & multi-stranded
fre arms) fastened to a single stranded frame th r eai^^MéMrM and the stem). One arm of the R Y consisted of
of’ the RY
several intertwined threads at R-F^;s^i|f^M wStf; thicker than any oiJ^^^^re#di’||^^M the wllr The thfe^â^^ -of the RY
the range of thickness (3 /x – 9 aO found for radii at
SiS-lj§|
Does the stre^lfi-èf a thread increase as its thickness increases? This $0jÈA 1 ikely because, generally, thread thickness corresponded
^ ^ ÉJLDPorting function of the
thread •> *-b°”W- large a portion of the web would collapse if it broke). Framgf^ were thickest; radiÿfrom the trapping zone and at the RY were next; and radii and NS’s from the strengthening zone, PS’s, and SS’s were thinnest.
Drag-line Attachment Disks:
Whenever the spider moves about outside of its web, it continually plays out a drag-line which it periodically fastens to the substrate. The ampullate gland which is involved in the production of the scaffolding of the web (frames, radii, NS’s, and PS’s) is also involved
20
Psyche
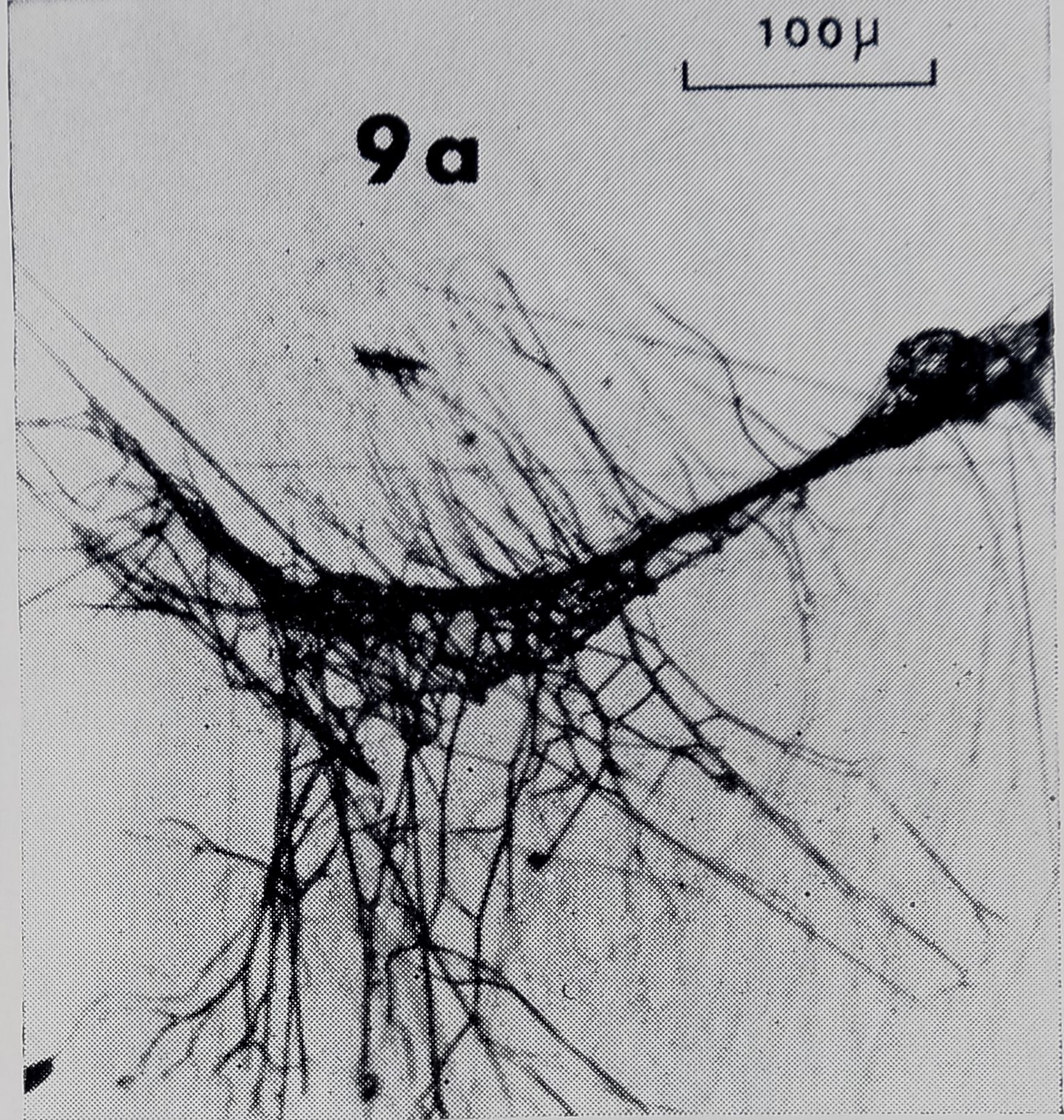
[March-J unein producing the dragline (Peakall, 1969). We examined two dragline attachment disks (Fig. 9) which were obtained by having spiders drop from glass microscope slides. Although these are not connections from the web, it would seem likely that mooring thread to non-thread substrate connections are closely related, if not equivalent, to drag-line attachment disks.
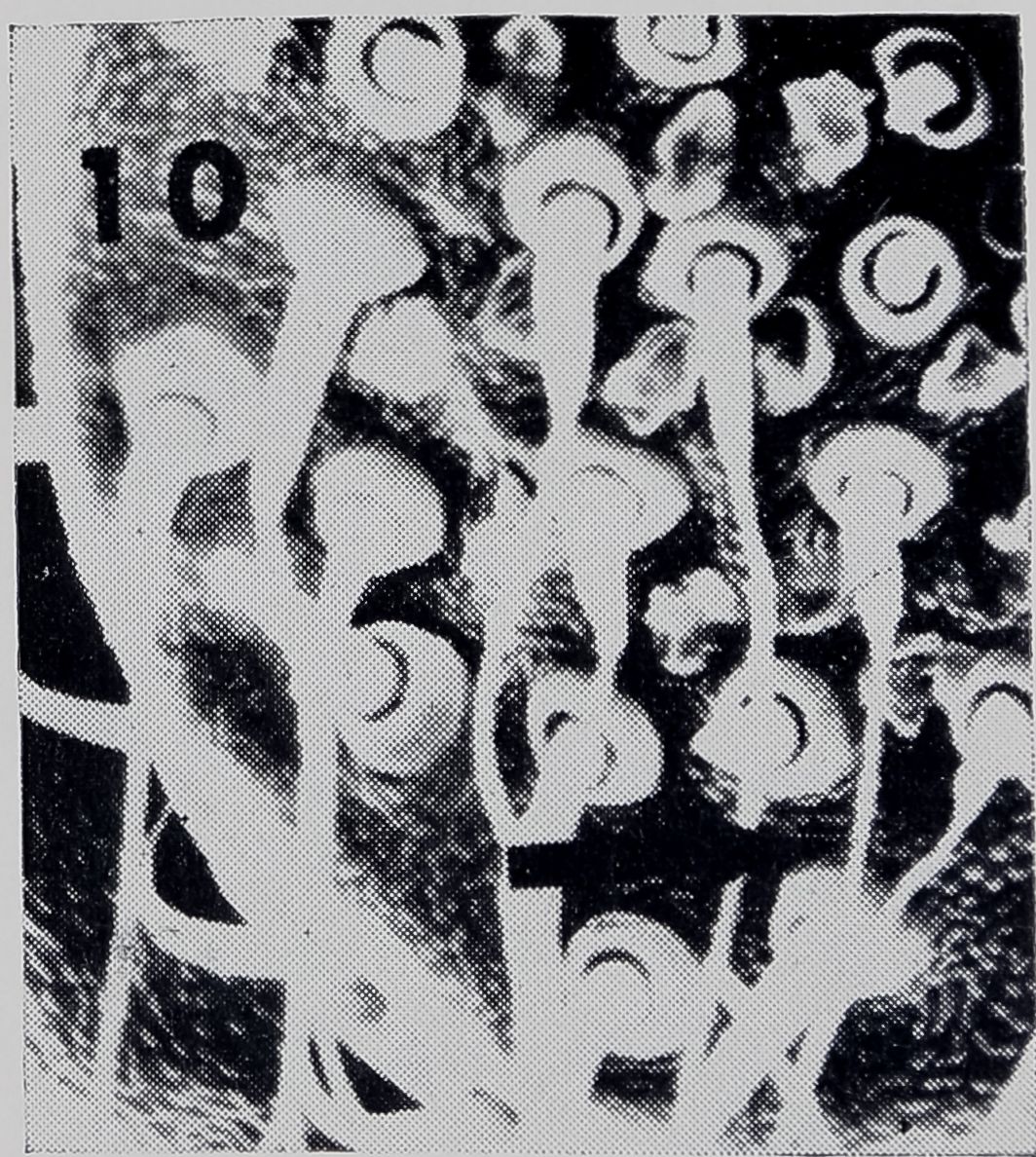
The production of the drag-line attachment disk is attributed to the piriform glands (Peakall, 1969). Threads between 0.5 /a and 1 fi thick can be seen emerging from the spools of the piriform glands in Fig. 10. The thinnest strands which could be resolved at the attachment disks were in this range of thickness (Fig. 9b). This thickness also corresponded to the thinnest strands of connections from the web. In particular, R-F’s, FY’s, and some SS-R’s had many strands in this range of thickness. This raises the question of whether the piriform glands are involved in the formation of each of these types of connections. One might hypothesize that some thread connections in the web are equivalent to drag-line attachment
Table 6
Descriptive statistics for some structural features of frame Y-structures (FY’s). For each feature, 5 FY’s from 3 webs built by 3 different spiders were observed. FY’s were rather variable with respect to these features, as indicated by the coefficients of variation. Comparing this table with
| Table 5, note the similarities | between R-F5 | s and FY’s. | |
| Feature | Means | Coefficient of Variation | Range |
| Length of Sleeve | 74.8 n | 95 | 22 jU-l8o jx |
| Number of Strands at Junction | 15.6 | 60 | 9-32 |
| Thickness of Stem of Y | 10.4 [x | 24 | 7 |
| Thickness of Single Stranded Arm of Y | 00
00 |
19 | 7 J®n IO |A |
Fig. 5. Thread connections from hub. a. Notice loose thread (A), connections resembling NS-R’s (B) (Fig. 3), and compound connections (C). b. Junctions of unusual shapes. Also note globules (G), which resemble those on SS’s in Fig. 2.
Fig. 6. Typical radius to frame connection. Radius, consisting of numerous strands, runs vertical; frame, horizontal. Frame can be seen to have sleeves at one side of radius and to consist of two strands at other side.
Fig. 7. Typical frame Y-structure. Multi-strand frame fastened to single 9trand frame.
197 n Jackson — Web of Araneus 27




Psyche
[March-June28

pig. 8. Radial Y structure. Sleeves can be seen on stem and one arm of Y. Note that beyond sleeve on stem, radius is split into several strands.
Fig. 9. a. Drag-line attachment disk with numerous strands of thread, b. Higher magnification. Note globules (G).
Fig. 10. Scanning electron micrograph of anterior spinneret showing numerous spools which open from piriform glands. Arrow indicates thread between 0.5 ^—1 H in diameter, coming from spool .1200X* Photomicrograph courtesy of Rainer Foelix, N. C. Department of Mental Health, Raleigh, N. C.



1971]
Jackson — Web of raneus
29
disks except that each thread is fastened to another thread rather than to a non-thread substrate. However; the number of strands (over 100, counting only those which touch the drag-line) at dragline attachment disks :(Figk 94} greatly exceeded the number at any connection the Also..globules %wyith^frh the exception
«é one PS-R, #ere ” dün the minute strands of con-
neCttefis frèm^ra^Webt, werfe- fo»®1‘ on soffit thick
strands |jfpPË<Ëne involved
fhë disks? What glands produce them? –
what
IMïf&fhin fhë ^Mndf,r’ the PeakSBHWWF*1
wStfM V w^e^^MBiiwP^^WWr taken under polarized light, of silk
gland which indicate that the silk -■ ét‘ the ^
tion into a solid is
located ins id e the spinner et an d 1f>rt for the ampullate
•piilraM
One hypothesis for junction formation is that^&^^^^^JtÉntiM| substance which’ |tite^#tAÆi^M»!^^he-f. As the. spinnerets touch AAfehyitt^^^^^^thread will be fastened, -the
then
connection to be with
At Material
from the aggregate glands. This conclusion is supported by two observ#ijpàf j globules
on th^ “SS ‘hu^^HplMdtton remained secure. 2)
Artificial sticky spiral to frame connections and IftR’s were produced bÿ placing a- êëftme or radius over a Permount basin, then placing a SS from the same web across it. Never securely fastened, the SS at these connections could always be moved in any direction on the frame or radius. SS’s which cross but are not fastened to radii are common in the horizontal web of Uloborus divcrsus (Eber-hard, 1969), and we found them in webs of A. diadematus after the spider had ben administered a central stimulant ( dextro-ampheta-
Psyche
[March-June30
mine sulfate, 100 mg/kg^ However, they do not normally occur in the vertical web of A. diadematus.
Possibly the material from the piriform glands is a cement. Does it cement the drag-line to the substrate through fusing to both, or are the threads ff@t$i the glands themselves cemented at one
end to the dragline and at the other to the substrate?
Stretching naay be responsifet^^^^^^ffication ©£ through
causing filaments to slide past one another, assuming positions which allow f®r a greater degree- of cross linkage (Lucas and Rudall,
1968 ). Another hypothesis ^yiunction formation is that flfS$ silk for the new thread fuses ^ith the old thread as it solidifiÿf., As the spinnerets sit on the some sJliid ^1% dfepfifts from the
spigot ol^fhé;>â^npullate gland and spools of any
other gland invc^^^i.. Solidification does not Occur becault fheve is insufficient aftefching of the mate||jjf. When thife lifted and sufficient stretching new* silkr solidifiesj gmd. is
fused to the old thread.
, information will be necessary before we- ^ahighoose between
these-hyp^tl®®^, Perhaps different çngchanisms: occur at different types‘ tmy .become Sp^isary to somttlfr
modilfear ■ combine these hypotheses.
’SUMMARY AND CONCLUSION^®
The fine slmwe at thread cdwnections ffl/ tpte gftjtb diadematus was examined by photomicroscopy. ^ffl?râcteristjc structures were discovered for each connection typé including gâseSfîfefce of ^S|#s’-bn lengths of junctions, Va^top|^Babers of
and different distributions of globules. Possibly and origins of structures were Thefe was of
two hypbtheies related tô ®bnne^t^p;?formatioh-: fojpHferteds are cementedijfogeth,er, and b) new thread,while in a semi-solid state, fuses to ioM thread.
This survey is only a modest beginning in the direction (Studying the fine structure of thread connections. With a more extensive and systematic investigate, tigor*^,. statistical comparisons could be made of different types of connections and of the same type of connection from different regions of the web. Structure could be compared for spiders of different age, weight, and species. Viewing the fine structure of connections has led to many interesting questions. Some were mentioned in this paper; others have probably occurred to the reader. Hopefully, these will be pursued further in future studies.
1^71] Jackson — Weh of Araneus 31
References Cited
Cowstock, J. H.
1940. The Spider Book. Comstock, Ithaca, N. Y.
DeWilde, J.
1943. Some physical properties of the spinning threads of Aranea diademata. Arch. nêerl. Physiol. 27: 118-132.
Eberhard, W.
1969. Computer simulator of orb-web construction. Amer. Zool. 9: 229-238.
Friedrich, V. L. and R. M. Langer.
1969. Fine structure of cribellate spider silk. Amer. Zool. 9: 91-96.
Jacobi-Kleemann, M.
1953. Uber die Lokomotion der Kreuzspinne Aranea diadema beim Netzbau (nach Filmanalysen). Z. vergl. Physiol., 34: 606-654.
Kullmann, E.
1957. Beobachtung des Netzbaues und Beitrâge zur Biologie von Cyrtophora citricola Forskal (Araneae, Araneidae). Zool. Jahrb. Aht. Syst. 86: 181-220.
Lehmensick, R. and E. Kullmann.
1957. Uber den Feinbau der Spinnfâden. In F. S. Sjostrand and J. Rhodin (ed.), Electron microscopy: Proc. of the Stockholm Conference, Sept. 1956. Academic Press, Inc., New York.
Lucas, F. and K. M. Rudall.
1968. Extracellular fibrous proteins: The silks. Comprehensive Bio-chem. 26B: 475-558.
McCook, H.
1889. American Spiders and their Spinning work. Vol. I, McCook, Academy of Natural Science, Philadelphia.
Nielsen, R.
1931. The Biology of Spiders. Vol. II, Levin and Munksgaard, Copenhagen.
Peakall, D. B.
1969. Synthesis of silk, mechanism and location. Amer. Zool. 9: 71-80.
Peters, H. M.
1939. Uber das Kreuzspinnennetz und seine Problème. Naturvoissen-schaften, 47: 776-786.
Savory, T. H.
1952, The Spider’s IVeh. Frederick Warne and Co., Ltd., London.
Wilson, R. S.
1969. Control of drag-line spinning in certain spiders. Amer. Zool. 9: 103-111.
Witt, P. N., C. F. Reed and D. B. Peakall.
1968. A Spider’s Web. Springer Verlag, Berlin.


