Warning: Trying to access array offset on value of type null in /home/elisew5/public_html/drpeterwitt.com/wp-content/plugins/contextual-related-posts/includes/content.php on line 49
Images Collection



View this article in Search Friendly Plain Text
NOTE: This plain text article interpretation has been digitally created by OCR software to estimate the article text, to help both users and search engines find relevant article content. To read the actual article text, view or download the PDF above.
CHANGES IN ORB WEBS OF SPIDERS DURING GROWTH (ARANEUS DIADEMATUS CLERCK AND NEOSCONA VERTEBRAT A McCOOK)
by
PETER N. WITT and RICARDA BAUM
ReprintedfromBEHAVIOUR Vol. XVI, 3-4

LEIDEN E. J. BRILL 1960
CHANGES IN ORB WEBS OF SPIDERS DURING GROWTH (ARANEUS DIADEMATUS CLERCK AND NEOSCONA VERTEBRAT A McCOOK) y)
by
PETER N. WITT and RICARDA BAUM Dept, of Pharmacology, State University of New York, Upstate Medical Center,
Syracuse, N. Y.)
(With 5 Figures)
(Reo. 26-IV-1960)
The web-building pattern of orb webs is characteristic for each species of spider, but within the same species there is considerable variation from one web to another. In working with orb-webs for many years it became apparent that each age group of spiders had its characteristic webs. We were finally able to tell the approximate age of an individual Araneus diadematus by looking at its web. There was, however, some difficulty in explaining on what such judgment was based. We believed at first that the web of an old big spider was always larger than that of a young individual. This is true if we go into extremes and compare the first web of a tiny spiderling with that of a grown-up animal. But the web of a 3 months old spider was not middle-sized in spite of the fact thaf\the animal was in the middle of its life. In the measurements reported below we have tried to establish a quantitative basis for the empirical observation that orb webs look different at different times of a spider’s life.
Part of the reason for our studies was our interest in web building as a test for drug effects (Witt, 1956). If influences which change web patterns of spiders are’ to be studied, it is essential to know the normal web. Changes in orb webs brought about by a drug can be identified only if the web which would normally have been built by a particular spider on the following day can be predicted. It is, therefore, believed to be of value to study all factors affecting web building including age. Several zoologists (Peters,
1) This investigation was supported by a research grant B-1794 from the National Institutes of Health, Public Health Service.
2) We are grateful to Dr Willis J. Gertsch of the American Museum of Natural History for identification of the spider species.
P. N. WITT & R. BAUM
310
1953; Petrusewiczowa, 1938) have discussed whether changes in web patterns during the growth of spiders are dependent upon changes in the growing spider’s body proportions. The appearance of a free sector in the adult Zilla web independent of previous practice has been used as an argument in favor of the theory that web patterns are innate. But this does not prove that other parameters in web growth are independent of the animal’s body measurements. Such considerations induced us to include in our report the results of measurements of the length of the first legs of spiders and their body weight.
Several authors have published results of their observations on webs of young and adult spiders of different species. These results are contradictory. Wiehle (1927) counted more radii ^1 the webs of young spiders than in those of adults while Koenig (1951) found less radii in the small webs of his young Araneus diadematus than in the webs of grown-up individuals. Savory (1952) observed a slight decrease in the number of radii during four summer months in outdoor Araneus diadematus webs but points out that there was considerable variation. Tilquin (i942)-Bwho counted radii in a great number of webs of Argiope bruennichi which he kept indoors— related them to molting periods. He describes a slight increase in average number of radii (from 22 to 29) around the third molting. For the rest of the spider’s life the average number stayed about even. His numbers varied between 10 and 40 radii in the webs of young spiders as compared to between 19 and 41 in adult webs. Peters (1953) measured mesh size between two radii and two turns of the provisional spiral, in young and adult Araneus diadematus webs. He found that there was a change in the ratio of body size to mesh size during the lifetime of a spider. This led him to the conclusion that body size does not determine mesh size. Mayer (1953) tried to establish a relationship between body weight and mesh size. One of her Araneus diadematus built one comparatively wide-meshed web with a low number of radii after a weight of 72 mg. had been attached to its body. But there is still some doubt whether the wide meshes were a function of the weight because the same spider with a smaller (30 mg.) weight attached built one even wider web.
All authors with the exception of Tilquin state or seem to imply that webs grow gradually in size as well as change their proportions uniformly during a spider’s lifetime. Not one, however, reports measurements which would support such an assumption. If it were possible to show that changes in webs are not gradual, the discrepancies in observations could be explained on the basis that observers watched the webs at different periods
CHANGES IN ORB WEBS OF SPIDERS DURING GROWTH 311
of the spider’s life. In some instances a low number of observations may also explain unusual results. It was, therefore, deemed imperative to keep as many spiders as possible under controlled conditions in the laboratory during their growth and observe carefully their webs over short intervals of time. The drawbacks of observations in the laboratory and the possibility of transferring results so obtained to outdoor webs will be discussed later.
METHODS
An egg cocoon of Araneus diadematus Cl was obtained outdoors and placed in a glass jar in the laboratory. On the 22nd of May, 1959, the first spider-lings were observed on the outside of the cocoon. All animals were left together in the paper covered jar until they started to attack each other. This was assumed to be the beginning of the period in which they would start to build individual orb webs. Each spider was then transferred to an individual glass jar and supplied with a 1.5 by 2 inch cardboard frame. The first webs were so delicate that it was difficult to remove them from the jar without damage. Web photographs were obtained as early as the second of June. These photographs represent either first or very early webs of each spiderling.
In August, 1958, several egg cocoons of Neoscona vertebrata McCook were obtained from spiders in the laboratory. Each cocoon was placed in a glass jar. By the end of August spiderlings had begun to emerge from all of them. They were treated as described above for Araneus diadematus. The earliest web photograph was obtained on the 19th of August.
As soon as it had started web building, each little spider was fed twice a week with Drosophila flies. It received water from the tip of a needle attached to a syringe. After 2 to 3 weeks the spiders were transferred into larger jars containing 3.5 by 7.5 inch frames, and the jar was covered with wire mesh screening. Six weeks after hatching each spider was put into a 20 by 20 inch wooden frame, 3 inches deep, which had glass sliding doors in front and in the back. In these boxes they received one to two house flies two times a week and water from a syringe. The spiders would pull the drop of water from the needle and drink it in 5 to 10 minutes. During the summer months temperature and light were kept close to outdoor conditions by means of open windows. As days began to get shorter a lamp with a timing device was installed which turned the light on at 5 A.M. and turned it off at 10 P.M. When outside temperatures dropped in autumn an automatic temperature regulation with a programmer was installed. This would permit us to raise temperatures in the early morning hours steeply from 10 to 25o C in 2 hours and cause a drop in the
Behaviour XVI
20
P. N. WITT & R. BAUM
312
evening. It was assumed that this corresponded roughly to light and temperature conditions on a summer day outdoors.
For web photography the glass doors were removed from the boxes. The webs were sprayed with white glossy Krylon spray paint. They were then placed in front of a wooden box which was painted black inside, and an even illumination of the webs was provided through flood lights placed on both sides. Each web together with a 20 mm measure was photographed on 35 mm. Kodak microfile film with a Contaflex IV camera. Special care

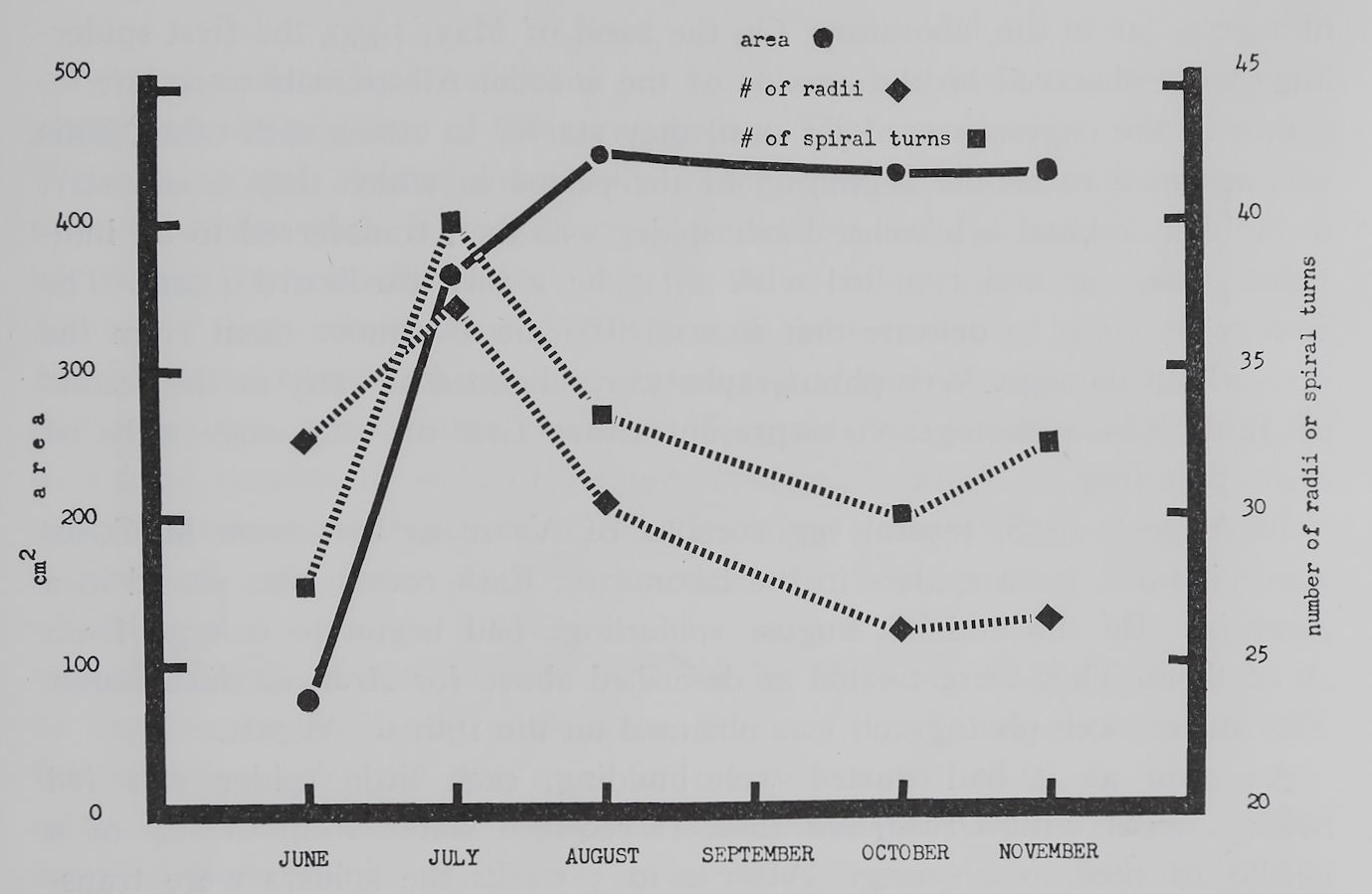
Fig. i. Average web size, number of radii, and number of spiral turns of webs of 14 individuals Araneus diadematus Cl. hatched from one egg cocoon on the 22nd of May, 1959. The single points represent 5 webs in June, 27 in July, 30 in August, 21 in October, and 20 in November. There was a significant change (P < 0.001) in web size from June to July and from July to August. The number of radii changed (l’Étil slightly from June to July and significantly (P < 0.01) from July to October. The number of spiral turns, which was counted in the lower part of the webs, increased significantly (P = 0.008) from June to July and decreased (P = 0.0004)
from July to October.
was taken that the film was parallel to the web so that distortions were avoided. Each web was destroyed after the photograph was taken, and the spider built a new web, usually during the following night.
To establish the size of a web its photograph was projected to the original size with the help of the 20 mm scale. The size of the catching area was
CHANGES IN ORB WEBS OF SPIDERS DURING GROWTH
3*3
determined by following the outermost spiral thread with a planimeter. The means of three such measurements per web was regarded as sufficiently accurate. Furthermore, the distance of the hub to the upper outmost spiral thread, lower outmost spiral thread, right and left was measured and called north, south, east and west respectively. Spiral turns were counted along one radius below the hub. Regularity of angles was measured as described by Witt (1956).
RESULTS
Figure i shows that there was no uniform steady increase in web size during the lifetime of Araneus diadematus. A fast increase in the catching
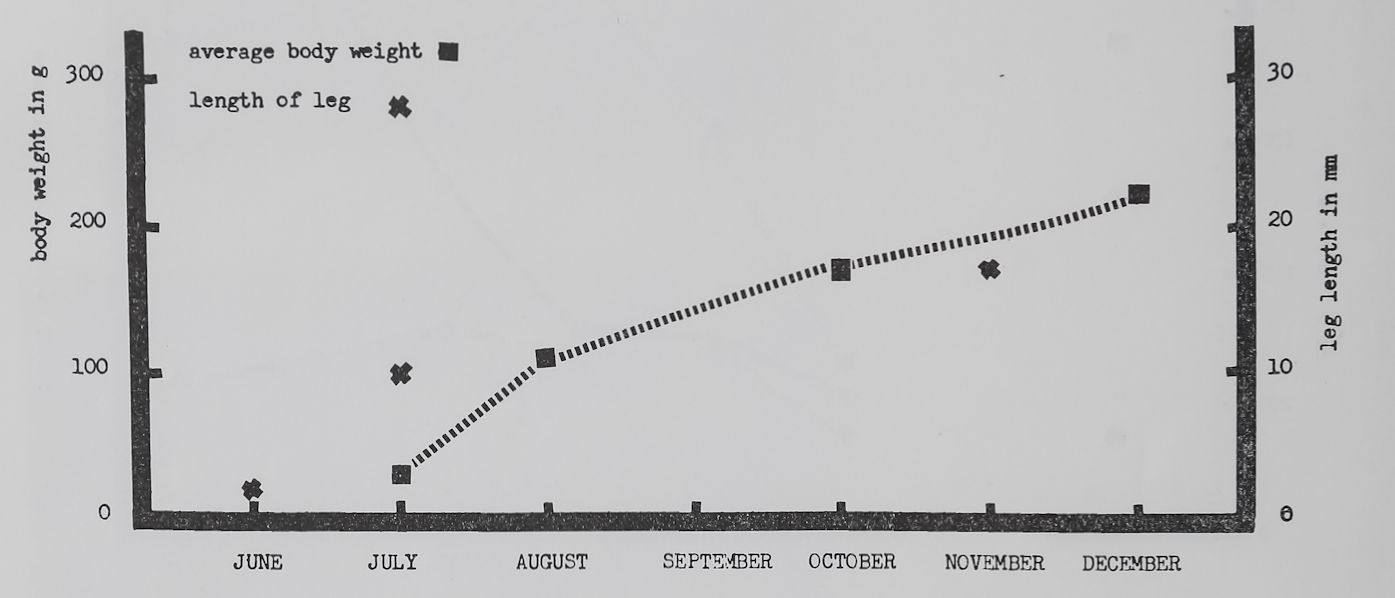
Fig. 3. Average increase H body weight of those spiders of which figure 1 shows the web measurements. Twenty-three weighings were made in July, 17 in August, 9 in October, and 8 in December. There was a statistically significant (P < 0.01) increase in body weight from eacOTpoint to the next. The crosses indicate length of first legs measured from femur to claws of the shed skins of one individual.
area (67%) from June to July was followed by a slower increase (17%) from July to August. For the next three months the size of the catching area did not change significantly. The number of radii and spiral turns changed apparently in a pattern different from change in size, but both changed in the same way. A maximum number of radii and spiral turns was observed in the not yet full sized webs built in July. The smallest area unit in the web which is limited by two neighboring radii and spiral turns can be called a mesh. Compared to their size the webs of the two month old spiders had a minimum mesh size. Later on the meshes of the webs became increasingly larger. The changes of the webs of our growing Araneus diadematus can be described as follows: during the first month

P. N. WITT & R. BAUM
314
of their life the spiders built small webs closely knit; in the second month larger webs with small mesh size were observed; large webs with increasingly big meshes were characteristic for older spiders. Characteristic examples of webs built by one spider in June, July and November of the same year are shown in figure 2.
Such observations become more meaningful if we compare the changes in web patterns with those in body weight and leg length (fig. 3). Leg length

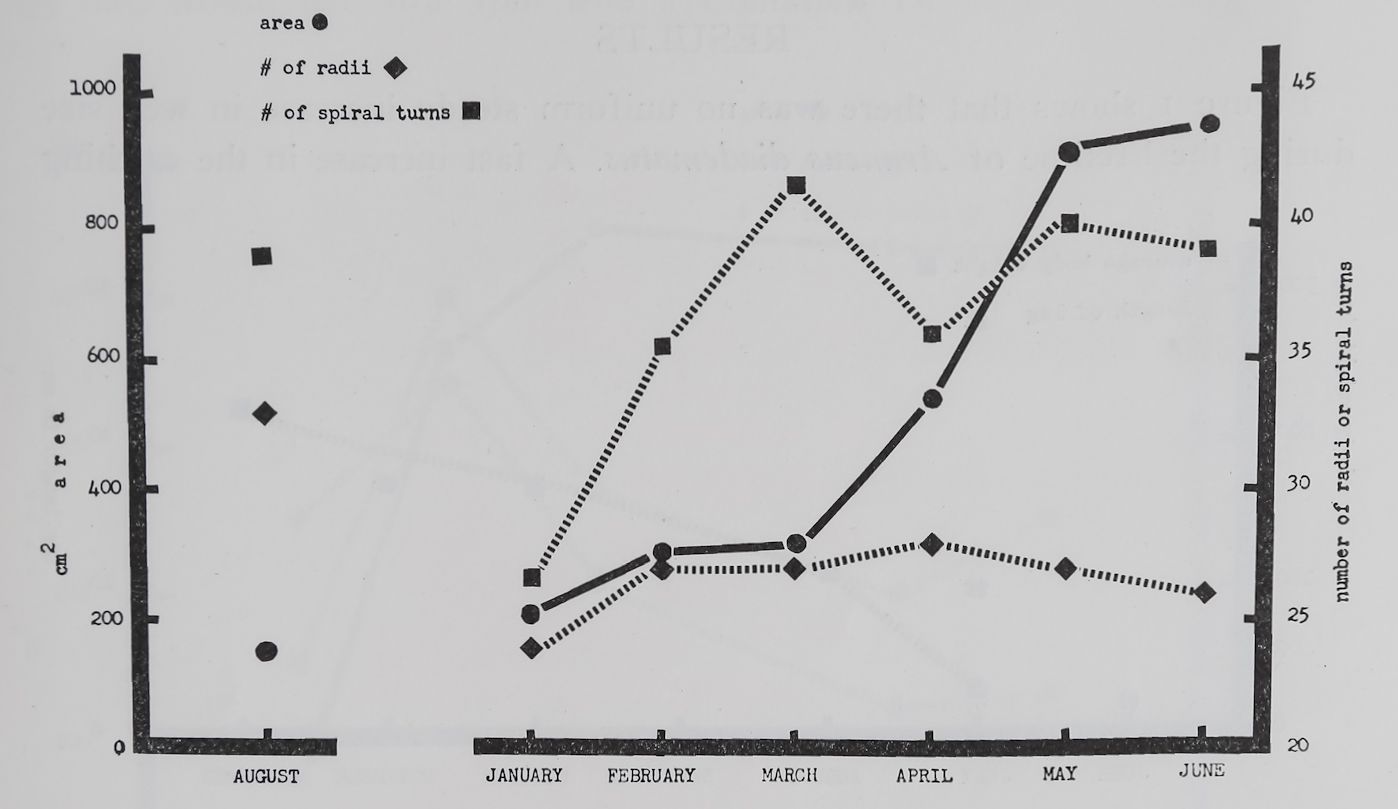
Fig. 4. Average web size, number of radii, and number of spiral turns in webs of 6 individuals Neoscona vertebrata McCook which hatched from 5 different egg cocoons in August 1958. The points represent measurements from 2 webs in August, 19 in January, 6 in February,, 3 in March, 13 in April, 17 in May, and 6 in Juñe. There was a significant increase (P < 0.01) in web size from March to April and May. There was no detectable change in the number of radii. A significant increase (P <[ 0.001) in the number of spiral turns was observed between January and March, and no other significant change. There is no statistical evaluation of the August data because of the low
number of observations.
increased rapidly (47%) during the first month and more slowly later (about 10% per month in four more months) in our Araneus diadematus. Body weight, however, increased more uniformly. This resulted in relatively light spiders with long legs in July as compared to heavier long legged spiders in November. Such a mode of growing can easily be confirmed in outdoor observations of female Araneus.
The pattern of changes in webs of growing Neoscona vertebrata was different from that of Araneus diadematus. This becomes apparent from figure

Fig. 2. Three webs of one individual Araneus diadematus Cl. built A) in June, B) in July, and C) in November. The upper part of B has been slightly distorted by the running spider. The black line indicates 20 mm in the original webs. The three webs represent some of the characteristic changes which occurred during the growth of the spiders. Observe the small area of web A as compared to B’ and C, and the small mesh size of A and B as compared to C.

Fig. 5. Three webs of different individuals of Neoscona vertebrata McCook which were born at the same time. Web A was built in August 1958 (and is slightly distorted), web B in February 1959, and web C in May 1959. The black line indicates 20 mm in the original webs. Observe the increase in size from web A to B and from B to C and the characteristic changes in mesh size.
CHANGES IN ORB WEBS OF SPIDERS DURING GROWTH 315
4. While catching areas did not grow much in size between August and March (4%) they increased rapidly thereafter to two and more times the March size. The final size was about 2)4 times that of full grown Araneus diadematus webs. The number of radii stayed about the same after an initial decrease from August to January, multiplied later, and showed the highest figure 2 months before the steep expansion in size started (March). From March increasingly larger webs with no change in number of radii or spiral turns were observed. Thus Neoscona, too, demontrates an increase in mesh size with increasing age. Figure 5 illustrates such observations in Neoscona webs from 3 individuals in August, 1958, February and May 1959.
The average body weight of Neoscona vertebrata in June (the last month of their lives) was 128 mg. The average leg length of 3 individuals for the same month was 20 mm. In comparing these figures with those of our full grown Araneus diadematus we find that the old Neoscona was considerably lighter and possessed significantly longer legs. No measurements of younger Neoscona have been obtained. There were no detectable changes in the relationship of length to width of web (vertical to horizontal diameter). We could not find any significant change in the symmetric position of the hub in the catching area (east over west) or in the regularity of angles between radii during the growth of our spiders. If any such changes do occur we may have missed them because of too small a sample size.
DISCUSSION
Neither bodies nor webs of either species of spider grew uniformly under our laboratory conditions. This leads us to conclude that there is no justification for making assumptions about webs at a certain age by interpolating from webs built before and after. This may explain some of the contradictory statements in the literature.
Our data confirm Peters’ observation that there is no simple relationship between leg length and mesh size in Araneus diadematus. There was in our webs, however, a relationship between leg length and web size and another equally simple relationship between body weight and mesh size. The young Araneus with relatively long legs and a light body built large webs with narrow meshes. When the spiders became heavier without showing a comparative increase in leg-length, their webs showed increasingly larger meshes. A similar relationship holds true in a comparison of the webs of heavy and relatively short legged full grown Araneus with those of the lighter long legged adult Neoscona. The webs of the latter were much larger in size and contained more spiral turns than those of Araneus. Some know-
3i6
P. N. WITT & R. BAUM
ledge of the spider’s body helps apparently in the prediction of its web pattern.
In the light of this knowledge the experiment of Mayer (1953) becomes particularly interesting. Her spider had to carry an added load for one day without a change in its leg length; this resulted in a web of which the meshes were wider than before. Such an observation is in agreement with the interpretation of our data given above. The effect of a drug which would make the spider build webs with larger mesh sizes (and we have made such observations) could be compared to the effect of an added weight to the body.
We do not assume, however, that body weight and ;kg length are enough to explain all the proportions of one web huilt on a particular day. Our graphs do not really reflect the day to day changes. As an extreme example of short range changes in the webs of a single spider we measured, on August 13, 1959, a web with a catching area of 542.4 cm2, on August 24, 1959 a web of 770.8 cm2, and on August 31, 1959, a,web with 229.2 cm2, area. This represents a 67% decrease in catching area in one week in the webs of one spider. Body weight and leg length are apparently only two factors out of a large number determining the web pattern. Their knowledge permits prediction of a general trend in proportion changes during a certain time of the spider’s life. Their usefulness is limited in the prediction of one web for a certain day, but is reflected significantly in the average of a large number of web measurements. For that reason we believe that in drug experiments, for example, a statistical comparison between a number of drug webs and control webs is necessary.
Apparently it becomes important to survey other factors which have influenced web building behavior of our spiders in order to establish the validity of the suggested correlation between animal size and web pattern. In another paper (Witt, i960) we have enumerated a number of factors which have been found to be of importance in web construction. Under our laboratory conditions, length of day, abundance of food, the daily rhythm of temperature changes, mechanical interference with web building, and gravity were kept as uniform as possible. The ample size of the wooden frames in which the spiders were kept can hardly have limited web size. There is a possibility that seasonal variations in barometric pressure and humidity produced some changes in web development. These variations could be responsible for some of the differences between Neoscona and Araneus, because the former were born in August and the latter in May. Such factors, however, could not have influenced individuals in each group differently because they all hatched from egg cocoons at the same time. We would expect changes similar to those observed by us in webs from
CHANGES IN ORB WEBS OF SPIDERS DURING GROWTH
3’7
spiders which grow up in other laboratories. But there might be a considerable difference in the rate of development of bodies and web patterns of spiders which grow up outdoors.
With our limited knowledge about spiders it is difficult to see any reason why a heavier animal should build a web with wider meshes or an animal with longer legs, a larger catching area. Such behavior may find an explanation through measurements of the thickness of the threads. Provided the volume of silk production does not increase at the same rate as the body weight in the growing spider and provided that the heavy spider makes the web out of thicker threads in order to make it sufficiently strong to hold it, the old spider may not have enough material for a large close-knit web. Measurements of thread diameters are planned. A narrow meshed web of large size seems useful for catching small and large insects in a wide area. In contrast a web with wider meshes would let most of the small insects pass through while it appears adequate to catch the large ones.
SUMMARY
Photographs of the webs of 14 individuals (Araneus diadematus Cl.) hatched from one egg cocoon and 6 individuals (Neoscona vertebrata McCook) were taken while the spiders grew up in the laboratory. The length of the first legs from femur to claws and the size of the catching area in the webs changed in a similar way: webs increased in size during the first two to three months of the spiders’ life as long as legs grew longer; after that both stayed about the same size. Body weight seemed related to mesh size. Light animals built webs with relatively many radii and spiral turns, heavy animals wide meshed webs. An explanation is proposed on the basis of the assumption that heavy animals have to build stronger threads with not more material than light spiders. Consequently they have less total length available l§r~t full sized web. A wide meshed web results.
REFERENCES
Koenig, M. (1951). Beiträge zur Kenntnis des Netzbaues orbiteier Spinnen. — Z. Tierpsychol. 8, p. 463.
Mayer, G. (1953). Untersuchungen über Herstellung und Struktur des Radnetzes von Arañea diademata und Zilla x-notata mit besonderer Berücksichtigung des Unterschiedes von Jugend- und Altersnetzen. H Z. Tierpsychol. 9, p. 337.
Peters, H. M. (1953). Weitere Untersuchungen über den strukturellen Aufbau des Radnetzes der Spinnen. — Z. Naturforsch. 8b, p. 355.
Petrusewiczowa, E. ‘(1938)- Beobachtungen über den Bau des Netzes der Kreuzspinne (Aranea diadema L.). — Trav. Inst. Univ. Wilno 13, (quoted after Mayer, 1953)-
Savory, T. H. (1952). The Spider’s Web-, London.
Tilquin, A. (1942). La Toile Géométrique des Araignées. — Paris, Presses Universi-taires.
Wiehle, PI. L. (1927). Beiträge zur Kenntnis des Radnetzbaues der Epeiriden, Tetra -gnathiden und Uloboriden. — Z. Morph. Oekol. Tiere 8, p. 468.
3l8 WITT & BAUM, CHANGES IN ORB WEBS OF SPIDERS DURING GROWTH
Witt, P. N. (1956). Die Wirkung von Substanzen auf den Netzbau der Spinne als biologischer Test. — Heidelberg.
– (i960). Web-Building. — Encyclopedia of Biological Sciences. New York, in print.
ZUSAMMENFASSUNG
Die Netze von 14 verschiedenen Kreuzspinnén (Araneus diadematus CI.), die alle aus einem Eikokon geschlüpft waren, und von 6 verschiedenen gleichalten Spinnen Neoscona vertebrata McCook wurden während des Heranwachsens der Tiere im Laboratorium regelmässig photographiert und ausgemessen. Ein Vergleich der Masse der Fangfläche in den Radnetzen und der Beinlänge der Spinnen schien eine Beziehung aufzuzeigen: während der ersten zwei bis drei Monate im Leben der Spinnen nahm Netzfläche und Beinlänge zu, während beide in den folgenden Monaten etwa gleich blieben. Das Körpergewicht hingegen zeigte Beziehungen zur Maschenweite im Netz (eine Masche ist begrenzt von 2 benachbarten Radien und zwei benachbarten Spiralfäden). Sowohl die ganz jungen als auch die älteren langbeinigen leichten Tiere bauten engmaschige Netze, während alte Spinnen mit schwerem Körper und im Verhältnis dazu kurzen Beinen weitmaschige Netze mit signifikant weniger Radien und Spiralumgängen bauten. Diese Beobachtungen, die durch die Versuche von G. Mayer mit beschwerten Spinnen gestützt werden, führen Verff. zu der Annahme, dass schwere Spinnen wohl’ dickere Netzfäden aus der gleichen Menge Material bauen, um ihr Gewicht zu halten. Sie haben deshalb weniger totale Fadenlänge zur Verfügung. So bauen sie ein weitermaschiges Netz, um mit weniger Faden die gleiche Fangfläche bedecken zu können. Es wird darauf hingewiesen, dass Spinnen unter der Einwirkung bestimmter Substanzen ähnlich weitermaschige Netze bauten.


