Warning: Trying to access array offset on value of type null in /home/elisew5/public_html/drpeterwitt.com/wp-content/plugins/contextual-related-posts/includes/content.php on line 49
Images Collection
Read OCR Digitized Article Text
NOTE: This plain text article interpretation has been digitally created by OCR software to estimate the article text, to help both users and search engines find relevant article content. To read the actual article text, view or download the PDF above.
Symp. zool. Soc. Lond. (1978) No. 42, 89-97
Comparative Feeding Data in Three Spider Species of Different Sociality: Araneus diadematus Cl., Mallos trivittatus (Banks) and Mallos grega/is (Simon)
P. N. WITT, MABEL B. SCARBORO and D. B. PEAKALL
NC Mental Health Research, Raleigh, North Carolina, USA Canadian Wildlife Service, Toxic-Chemical Section, Ottawa, Canada
SYNOPSIS
Patterns of feeding behavior in three species of spiders were measured, and interpreted as web-related survival strategies, possibly correlated with communal living of the species. Lively houseflies loaded with tritium-labelled glucose were placed under controlled conditions in the webs of hungry spiders, and time to attack and to feeding was measured: the two Mallos species attacked later, but started feeding at about the same time as Araneus diadematus. After 10, 30, 90 min or 4 and 24 hours, distribution of solution between spiders and flies was measured ill a liquid scintillation counter. Speed of feeding was correlated with body weight, but not with age in A. diadematus; both Mallos species fed significantly more slowly. In small colonies of M. gregalis individual animals ate widely varying amounts, increasing in number of drinkers and maximum amount drunk over time; always several animals did not drink. It is postulated that web (permanent-space vs temporary-orb) and silk type (cribellate vs sticky) largely determine attack behavior, while pattern of feeding is correlated with each spider species’ way of life and possibly with communal versus solitary living.
INTRODUCTION
Tolerance toward conspecifics, communal feeding behavior and gregarious lifestyle appear closely correlated in the predatory spiders (Kullmann, 1972): where centered orb-webs are the substrate for solitary living, spiders attack and mostly feed on any moving, weighty object, including conspecifics (Peters, 1933). Earlier in life, gregarious spiderlings of the same species tolerate each other on the irregular space web (Burch, 1977). In social spiders females, males and juveniles live together on one space web and attack and devour prey in groups. Other spiders apparently occupy in-between positions, where two or more animals can live on one web and feed on one prey (Jackson, in press). Frequently these behavioral differences go together with differences in body size, phylogenetic relationships and web structures. In our evaluations we tried to compare three species of different sociality in a relatively simple measure, i.e. quantity and pattern of food uptake. The feeding
P. N. WITT, MABEL B. SCARBORO and D, B. PEAKALL
90
results can be compared to various other behaviors in the hope of identifying some of the options which communal life presents for survival in specific situations. Thereby we extend measurements of Krafft (1965, 1971), who investigated questions of feeding in Agelena consociata Denis, another social spider. It is our goal eventually to measure the energy flow in a spider society, in comparison with energy data obtained on a solitary spider (Peakall 8c Witt, 1976).
METHODS
Of the three species spiders selected, one, Araneus diadematus Cl., lives alone in an orb-web which is renewed daily. It sits motionless for nearly 23 hours, except when prey -touches the web. The prey is attacked, bitten, wrapped in silk and transported to th0%ub ©f the- web. A few minutes kjtffEP the spider can be seen |b‘$tart sucking On the ffp, the feeding period lasted from that- moment until fbttiutes^ to several
hours later when We terminated $£ by removing the prey,, All animals were raised in the htborat©rydfr.Qm çfgç&gh|^btained from Upstate New York (Witt, I#7l). Raising animals from ,permitted u$ to take
them into ouf Experiment at .various designated stages of weight, (agefei
Mallos trivittatus (Banks) builds irregular, long-lasting sheet webs; the spiders ^ack aifit teisuipe pttf similarly’lit? A. diadematus, but without wrapping. Sometimes two spiders dbed together o^|one fly. These characteristics have led us to ctfnsider them to éilupy a-fWffee between the solitary A. diadematus and tftétcomniîînâl M. gregalis. All our animals
were adulüè ‘Which had been caught by R. R. Jackson more than six months earlier ii$’Portal, and kept in small plastfc containers.
; ‘Colonies of M. gregalis (Simon| w^fl^^^^^Mainly of adult females with an occasional male d0_ Juveniles live and hunt together on a sheet-covered space*web (see Burgess, 1976b Our colonies were descendants from 9 group of animals obtained more than two years earlier at Guadalajara, Mexico by J. W. Burgess. Several weeks before the tests, small colonies of three to 20 animals had been isolated in glass containers of lO-cm diameter and 13-cm height on wooden frames. Several days before each experiment, all le^Svers from feeding and debris were eliminated. It can be assumed that at the start of each experiment all spiders were hungry.
Radioactive glucose D»[f-3H(N)]-HOCHdCHOH)4CHO was diluted with cold glucose in water, so that five microliters would count close to 900 000 counts 10 min-1 in the scintillation counter. Five microliters of the solution were injected with a Hamilton microsyringe into the abdomen of Musca domestica. Flies which showed leakage or died were discarded. Ten minutes after the injection, lively radioactive flies were positioned with forceps into a web, and attack and start of feeding were
FEEDING IN SOCIAL SPIDERS
91
timed with a stopwatch. Spiders were given 10, 30, 90 min or 4 and 24 hours to feed.
At the end of the feeding period, the container with spider and prey was placed into a freezer (temperature —6°C) for one hour. Each animal was taken out with forceps, weighed and placed separately into 2 ml of protozol for digestion and homogenization. In long-term experiments silk was also digested. After addition of scintillation fluid and cooling, samples were counted several times and compared with a calibration curve derived from the same solutions without animals. Background counts were subtracted. As controls, several flies were injected and prepared for counting after 10 min without presentation to the spider. The sum of all counts from one experiment is called 100% ; it comprises the portion of glucose drunk by the spider and the portion left in the fly. Recovery was close to, but tttftly exactly
Autoradiographs were made of flies which were fixed ft) min after injection with labelled glucose, and of spiders which had fed for 90 min. Animals were fixed in 10% formaldehyde and subsequently embedded in wax. Ten-micron sections were coated with NTR-2 tracking emulsion and developed after three and eight days. Sections were stained lightly with Azure B and counts made of the number of exposed grains per unit area. Results are expressed as counts per «unit area per unit 0§ body weight. All values are averages of 100 Counts.
RESULTS
The period from the moment at which the fly Was placed ®n the web to the time of the first spider bite fall’s called attack tiifie. There was a highly significant difference between this time for A. diadematus (mean S*4® i) compared with either M. trivittatus (mean 20.8 min) or M. gregalis (mean 37″5 min); $towever, there was no significant difference between the two Mallos species. The solitary orb-web builder always attacked sooner than the communal spiders.
All three spider species would either transport or leave the prey after the first attack. In either case, the moment at which sucking started, several minutes later, could be identified clearly. The time between first attack and sucking varied widely from spider to spider, and we could find no significant differences between our three species.
Another interesting comparison between species can be made by measuring how much the animals drank in the first 90 min of feeding time (for M. gregalis the whole colony was treated as one individual, for A. diadematus we selected 81-day-old and for adult ani
mals): while M. gregalis drank an average of 25*9% of the fly, M. trivittatus drank 33*9% and A. diadematus drank significantly (p < 0.01) more than both, namely 67*5%.
92
P. N. WITT, MABEL B. SCARBORO and D. B. PEAKALL
There is a clear difference between the body weights of adult females of the three species: 14 M. trivittatus used in these measurements weighed a mean of 31*3 mg, with a range from 6*8 to 60*0 mg; (the lowest, a male, was disregarded in the graph); 35 female M. gregalis weighed a mean of 8’2 mg, with a range from 3*8 to 20*9 mg. Adult female A. diadematusvary widely in body weight, and because we had
raised them from egg cocoons m the laboratory, we knew their age and were able to measure their food uptake at different age (weight) periods: the youngest group of A. diadematus consisted of IS individuals, 31 days old, weighing a mean of 8.32 mg; the next consisted of 10 individuals, SI days old, weighing?®*? mg; nine spiders 105 days old weighed about 105 mg; at age 1$5 days we separated two groups from each other, the first with nine individuals, weighing a mean of 227*8 mg, the other consisting of six individuals, weighing a mean of 34* 1 mg. We can therefore compare amount drunk between species, and kjfone species (A, diadematus) ai different ages and body weights.
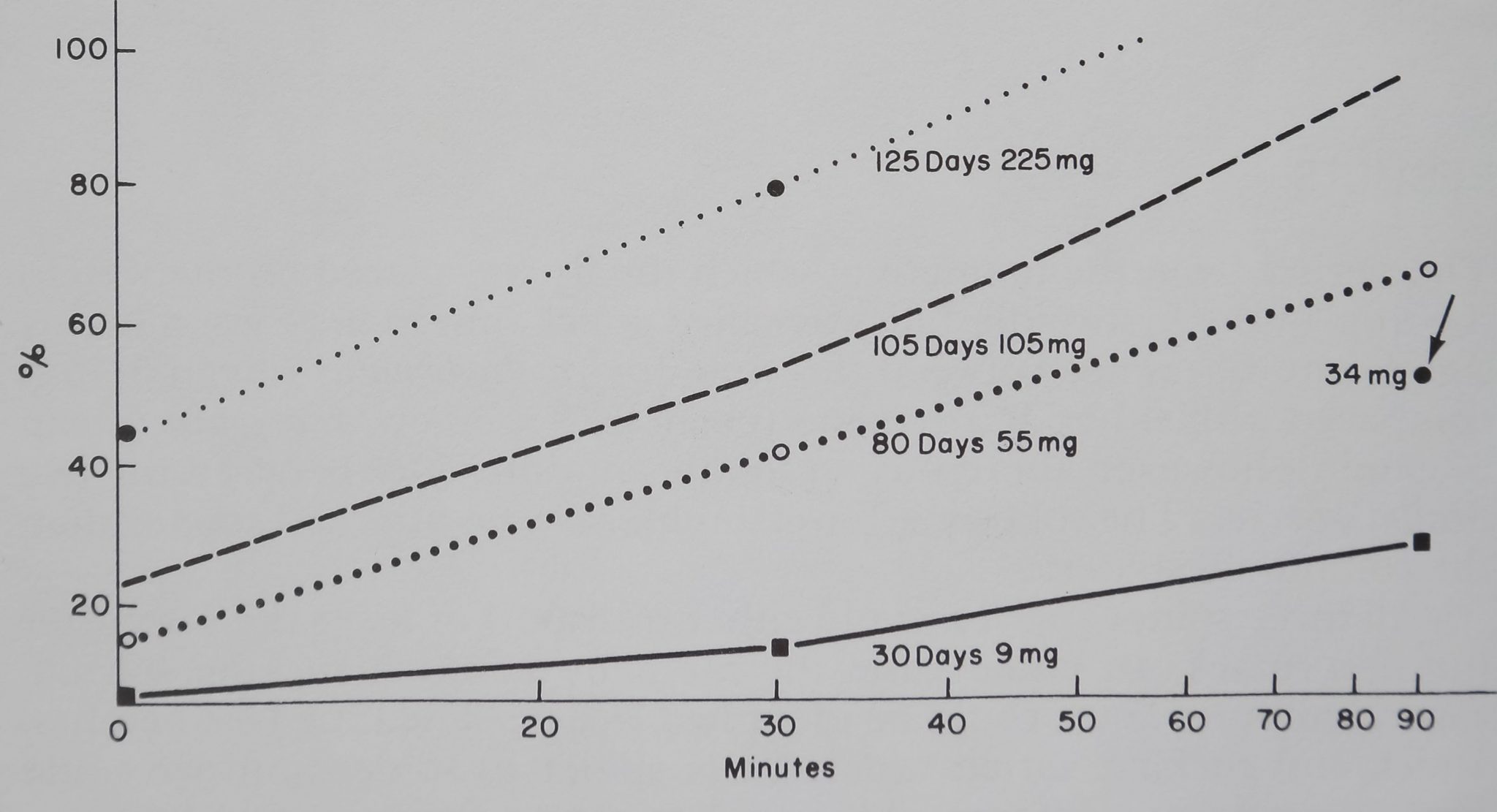
It has already been shown that A. diadematus attacked prey very; quickly,-in many experiments at the moment-jt touched the web. Thereafter they fait, wrapped and transported the prey to the hub and usually waited several minutes before starting to drink, presumably for the purpose of letting poison and gastric juices take effect* Figure 1

Fig, 1. Each point on the graph represents the mean of 2-6 measurements of radioactivity (ordinate) in Araneus diadematus spiders of four different age and five different weight (more exact weights in text) classes, at various times after sucking on the radioactive fly had started (abscissa). Note the increasing amount of food taken up by the spiders over time; the heavier the spider, the larger the amount she can drink in a given time. That it is weight and not age which determines quantity of food is shown by comparing the 30-min value for the oldest spider group at 225 mg (about 20 min later prey had been emptied and was discarded) with the 90-min value of spiders of the same age, but smaller site (marked by arrow).
FEEDING IN SOCIAL SPIDERS
93
shows the percentage of radioactive glucose in the spider, as compared with the relative amount left in the fly after various times and for different weight (age) groups. Since autoradiographs of three flies made 10 min after injection of label showed that radioactivity was fairly uniformly distributed in the abdomen, it seems safe to assume that the amount measured is representative of the amount of fly contents drunk. Fly weight varied with drinking time, being initially between 10 and 20 mg, reduced to 1-2 mg after longer sucking periods. In the long-term drinking experiments fly remnants were sometimes discarded by the spider onto a paper fastened below, and weight and counts were obtained from these. There can be hardly a doubt that A. diadematus drank in proportion to its own weight (size) and time it was left with the prey; age did not play a significant role.
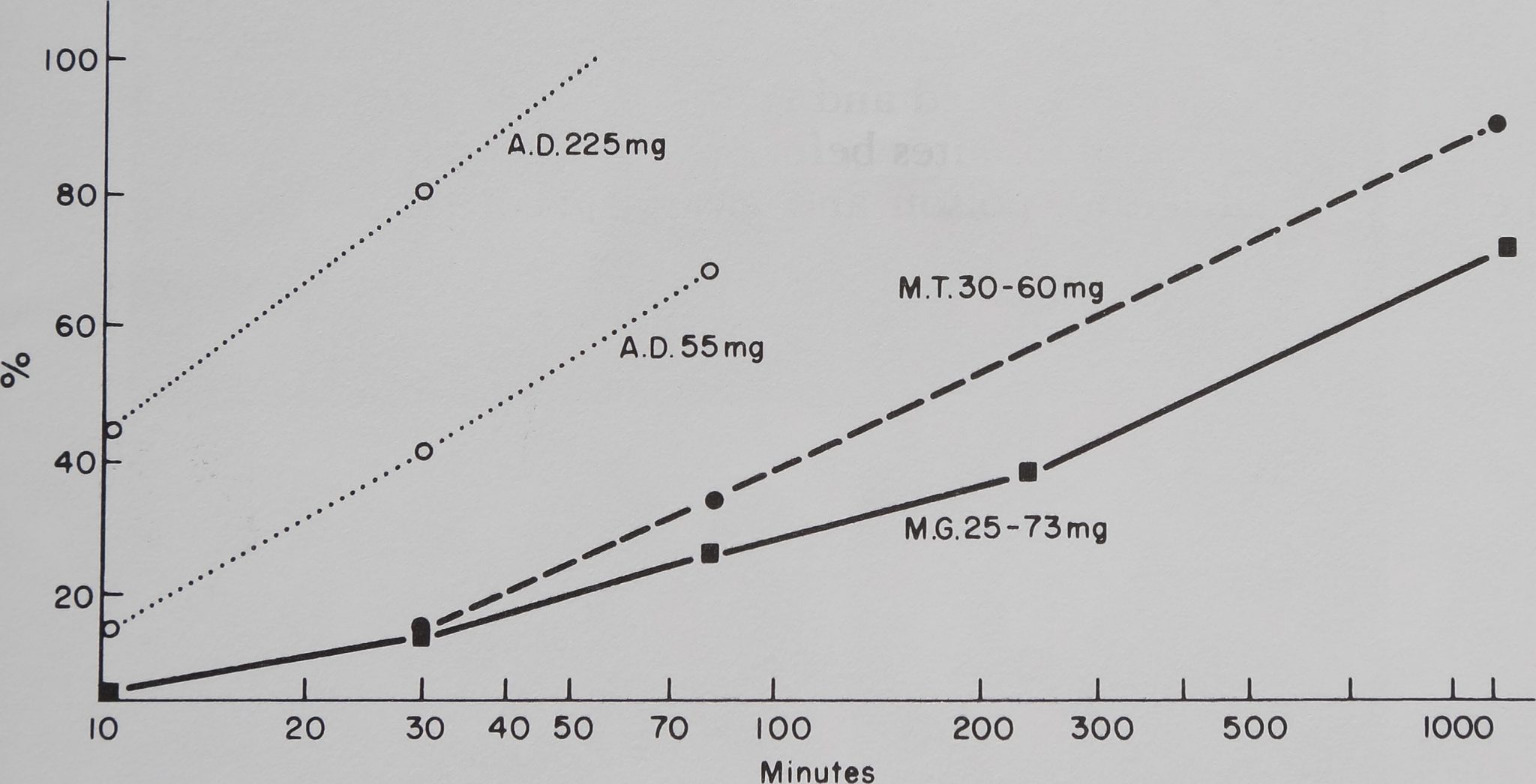
A comparison of the three species presents more problems: it can be done by age (mature females only) or by weight. Figure 2 explores both

Fig. 2. In this graph, in which feeding data for three spider species are compared, abscissa and ordinate are as in Fig. 1. Only the heaviest and one lighter group of Araneus diadematus (A.D.) are shown; the first represents adult females (as in the other two species), the second animals of weights comparable to the other species. For Mallos (M.G.) the sum of
the weights of all the individuals in one colony who had fed on the radioactive fly is listed. Note the significantly slower feeding pattern for the two Mallos species, as compared to A. diadematus; at 90 min M. trivittatus (M.T.) had consumed significantly (p < 0.01) more of
the fly than a colony of M. gregalis, but at 24 hours there was no significant difference.
possibilities. It shows that A. diadematus was always the fastest feeder, whether tested as juvenile in a comparable weight range to that of M. trivittatus or as adult. The solitary spider completely devoured and discarded a fly in one to three hours, while M. gregalis colonies took more than 24 hours, and M. trivittatus was in between. For a comparison of body size and feeding mode see Fig. 3.

FEEDING IN SOCIAL SPIDERS
95
In M. gregalisthere was an increase in the number of individuals
sucking on the fly over time, from an average of 4*5 animals in the first 10 min to 7*2 animals in 24 hours; however, in none of our small colonies, which consisted of animals sitting close together, had all individuals fed in one day. Because of high variation from spider to spider, we were unable to show that significantly more had been devoured per spider in 24 hours as compared to 10, 30 or 90 min. However, when single feeders could be identified, the maximum drunk by a spider increased from 4*15% of the solution in the first 10 min, over 30*1% in 90 min, to 74*6% in 24 hours, a remarkable amount of feeding in such a small animal. Body weight had increased to over 20 mg in one of the heavy feeders of M. gregalis.
The autoradiographs of A. diadematus and M. gregalis after 90 min of feeding showed ggtnsfderable radioactivity in the abdomen, except for the silk glands and muscle, which had levels close to background. The autoradiographs of M. gregalis showed considérable variations he. 81-370 counts mm 2 x body weight, but were than the two
A. diadematus, which had meah eounts of .481 and 637. The comparable values for the three flies were 810-1076. Tbe results agree well with those; of the scintillation counts and confirm the assumption that we measure distribution of food;
DISCUSSION
M our Observations WRÈfe made i^j^ laboratory^which has the aid vantage ©f controlling environmental conditions (e.g. being sure that animals have been kept without food several days before the measurement® and providing uniform pfif (adult M. filled for 1<0 min with a
known amount of radioactive glucose). It must be kept in mind, however, that there can be a grift amount of variation in the natural environment of the three species, and that colonies of M. gregalis can be very much larger than those utilized in this experiment. Despite that caution, it seems reasonable to believe that radioactive measurements in the spiders provide figures which are representative of general feeding patterns.
Our method did not permit us to measure individual drinking times in M. gregalis, nor can we say how each member of a colony would behave in repeated feedings; such observations are presently being made by S. Smith in this laboratory.
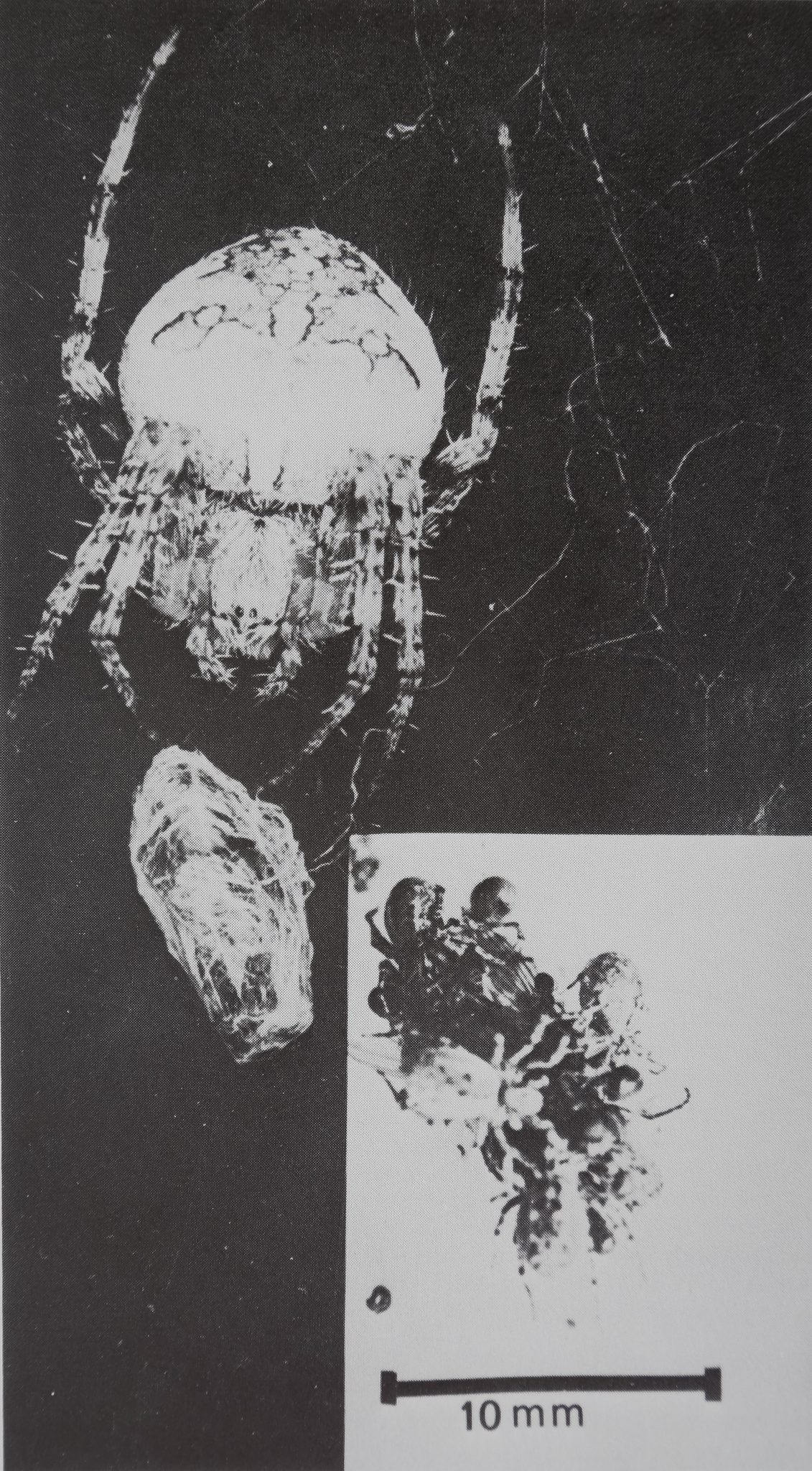
Fig, 3, An adult female Araneus diadematus (in the left upper corner) has wrapped a house fly and sits in the hub of its web, ready to feed. In the lower right corner several adult female Mallos gregalis sit on a house fly, starting to drink. Note the difference in size and number of spiders in the two parts of the photograph, which are enlarged to the same scale. Measurements and observations have shown that not all M gregalis which have subdued a fly drink from it afterwards.
96
P. N. WITT, MABEL B. SCARBORO and D. B. PEAKALL
What becomes apparent is the relatively slow way in which both Mallos species attack and devour their prey; it is as if the fly caught in the web was considered by Mallos to be a food source for an extended period of time, to be returned to perhaps at a later date, whenever other activities permit. In contrast, Araneus pounces quickly on the prey and sucks it out without interruption, the larger animals emptying the prey more quickly than the smaller. Comparison of the two types of webs comes to mind. The attack mode of the three spider species seems to be a function of their different webs. At least, the permanent, cribellate web enables the unhurried approach of the two Mallos spiders. Their prey is trapped securely in a dense structure. By contraMj the sparse, economical and open structure of the web of A. diadematus detains the struggling insect on sticky threads for perha$#~i9ajy a short time; the spider secures its prey quickly tty rushing out to bite and bind it.
Beyond that we would like tqispeculate- that the behavior patterns of A. diadematus and M. gregalis show a difference Wblriil is expressed in the spider’s whole living style: the “single-minded” attack of the solitary spider, her immediate and unswerving attention to «Ike securing and devouring at prey, is- one episode in m composed of sever*
all-or-none categories of action. The orb-web builder spends much time in immobility^jpunctuated bfiiUN^igly patterned activifrp There is a time to build a web, a time ro mate a®* most of wa^j When
feeding is to bt is transacted,
M. gregalis, and in some ®Ély M. trivittatus, appear to have ever-changing patterns of ot; le# activity; these are not distinct:
schedules devoted to single prèjèQ^ except * occasion demands. Attack and feeding practices are qansistent with this apparently more flexible behavior. The fly will be approached and devoured sometime, but its entanglement is not a signal to drop everything else. More observation® are needed, but we agree with Kullmann (1970) that “dem Zusam-menschluss (in Spinnen) liegen offensichtlitth i.. ernihrungsphysiolo-gische Vorteile zugrunde!”
ACKNOWLEDGEMENT
The experiments were in part supported by a grant from the National Science Foundation to P. N. Witt. Special thanks for help to R. Daniels and C. F. Reed.
REFERENCES
Burch, T. L. (1977). Early communal life of the solitary web-building spider (Araneus diadematus Cl,). Ph.D. Thesis, N. C. State University.
Burgess, J. W. (1976). Social spiders. Scient, Am. 2S4: 100-106.
FEEDING IN SOCIAL SPIDERS
97
Jackson, R. R. (In press). A comparative study of social organization and web characteristics of dictynid spiders. Revue Arachnol.
Krafft, B. (1965). Sur une possibilité d’échanges de substance entre les individus chez l’Araignée sociale Agelena consociata Denis. C.r. hebd. Séanc. Acad. Sri., Paris 260: 5376-5378.
Krafft, B. (1971). Contribution à la biologie et à l’éthologie d’Agelena consociata Denis (araignée sociale du Gabon). Biologia gabon. C7: 3-56.
Kullmann, E. J. (1970). Bemerkenswerte Konvergenzen im Verhalten cribel-later und ecribellater Spinnen. Freunde Kôlner Zoo. 4: 123-150.
Kullmann, E. J. (1972). Evolution of social behavior in spiders (Araneae: Eresi-dae and Theridiidae). Am. Zool. 12: 419-426.
Peakall, D. B. & Witt, P. N. (1976). The energy budget of an orb web-building spider. Comp. Biochem. Physiol. 54A: 187-190.
Peters, H. M. (1933). Weitere Untersuchungen über die Fanghandlung der Kreuzspinne (Epeira diademata CL). Z. vergl. Physiol. 19: 47-67.
Witt, P. N. (1971). Instructions for working with web-building spiders in the laboratory. Bioscience 21: 23-25.

 III. PREY AND PREDATORY BEHAVIOR<br><strong>PSYCHE</strong>")
 I: social organization and web characteristics<br><strong>Revue Arachnologique</strong></br>")