Warning: Trying to access array offset on value of type null in /home/elisew5/public_html/drpeterwitt.com/wp-content/plugins/contextual-related-posts/includes/content.php on line 49
Images Collection
View this article in Search Friendly Plain Text
NOTE: This plain text article interpretation has been digitally created by OCR software to estimate the article text, to help both users and search engines find relevant article content. To read the actual article text, view or download the PDF above.
Growth of the Orb Weaver,
Araneus diadematus,
and Correlation with Web
Measurements
BY J, SFNFORM}»
and
K. H.
Reprinted from Psyche, Yol. 80, Nos. 1-2, March-June 1973
pages if-lfi§ _ ;
GROWTH OF THE ORB WEAVER,
ARANEUS DIADEMATUS, AND
CORRELATION WITH WEB MEASUREMENTS*
By Jay Benforado and Kent H. Kistler
Division of Research, North Carolina Department of Mental Health
Raleigh, North Carolina iyfeii
iNTRODueytha^H
It is a well-known fact that within anypopulation of spiders of
similar age there tlfiftion ig^iftij dm of individual
spiders of the same species. In lifëïàtjire as eaiW’ m 1^90;,, McCook
has observed £bêàs< /and .* rëpèatefl (^ervati’Orîs ( Comstock,
th^phenomenon. Although ob-
servations are’^®j^n>t, explanaftiçjps Rria$ÉN&
feites differencetln feeding as a reilphJIo.pidifferential site, but the
mjdmgjîy^n messing-and to the authors’ knowledge
yb no#, el^^jed.’upon elsewhere. ThS^p^^wB explanatlffift lends
iteelf to Jjfarthif ;|ipp,|sfe of the factors contributing to the phenom-
enon
^ £aper mjlkoisolate so^e lof thé. Infers which
contrfl&Crte to ‘mjajHn’ • A ?ôr0ig \ diademwtMS O^rck (fa#
and to elaborate ^^&t:’fer-
taip» 0^®e§êffa^orfs able.
^^^Sespqnding with differential ’ size, in an. ^rb-weaver such ai
A rompus dïadematus,àift.£rent\a\ growth iVt$lso manifested ^.chang-
ing Æthens-ions o|^ihe That ]arf^:ÿfl^ÊÊÊti^. in dimensions
exist betwen |Ejj$ indi^Jlual Webs of spider* it also a well-known
fact. An artèmp|;.^^laip^é^ié o^^Œfactors influencing web
changes is also made.
Method
environment; mbe spiders usfit were from two cocoon® of Ara-
neus diademutiiSj obtained from Canastota, New York, which hatched
on April 26, 1972. From the time ^f hatching and throughout the
experiment, the spiders were kept in a room which was lighted 16
hours per day aand kefk cool during jthe eight dark hours by an air
conditioner. (See Witt, 1971^, ;
EARLY rearing: At the time of hatching, the offspring from each
* Manuscript received by the editor March 1,1973
90
1973]
Benforado Kistler — Araneus diadematus
9i
cocoon were placed in a separate rearing box. The spiderlings were
kept in these boxes, living on a communal web with a constant sup-
ply of loose gnats in the box, until they began to build individual
webs approximately three weeks after hatching. As each animal built
her first web she was removed from the rearing box and placed in
an individual glass tube, approximately 1X7 cm, with the ends
of the tube stoppered with cotton. From the time the animals were
placed in the tubes until onset of the experiment they were fed ap-
proximately 10-15 gnats per week, by placing the gnats in the tube
with the spiderling. The animals were watered by wetting the cot-
ton with water daily.
distribution : Seven weeks after hatching the two sets of spider-
lings were each separated into three equal groups by means of a
random numbers chart. No attempt was made to distribute males
and females evenly. Although the growth (body weight) of males
and females differs, it has been shown that the early growth of both
sexes is alike (Witt et al., 1972). Bepause of the short duration of
the experiment and the difficulty in identifying male spiders before
the last molt, distribution of males and females was left to chance.
At the time of initial grouping the two sets of spiderlings num-
bered twenty and thirty respectively. It was decided to feed each
of the three groups of each set according to a different feeding sched-
ule: one group every day, one group twice weekly, and one group
every ten days. Thus there were six groups, one for each set of
offspring on each feeding schedule. After one week of this procedure,
however, it was decided because of the small size of the groups to
reduce the number of schedules to two, and the middle schedule was
dropped and its members distributed randomly between the lighter
and heavier-fed groups.
Data for animals that died or escaped during the course of the
experiment were removed, so figures represent only animals observed
for the duration of the experiment.
weighing: Each spider in the heavy-fed groups was weighed once
a week, to o. 1 mg, while animals in the light-fed groups were
weighed on the day of feeding and the day after feeding.
web analysis: After eleven days of controlled differential feeding
in the tubes, the spiders were transferred to aluminum and glass
laboratory cages, 50 X 50 X 10 cm. At this time the animals were
eight weeks old. From this time on the spiders began to build webs.
Photographs of webs were taken daily and analyzed (Reed et al.,
1965). Daily records of web building were kept and the webs were
Psyche
[March-Jufii
Q2
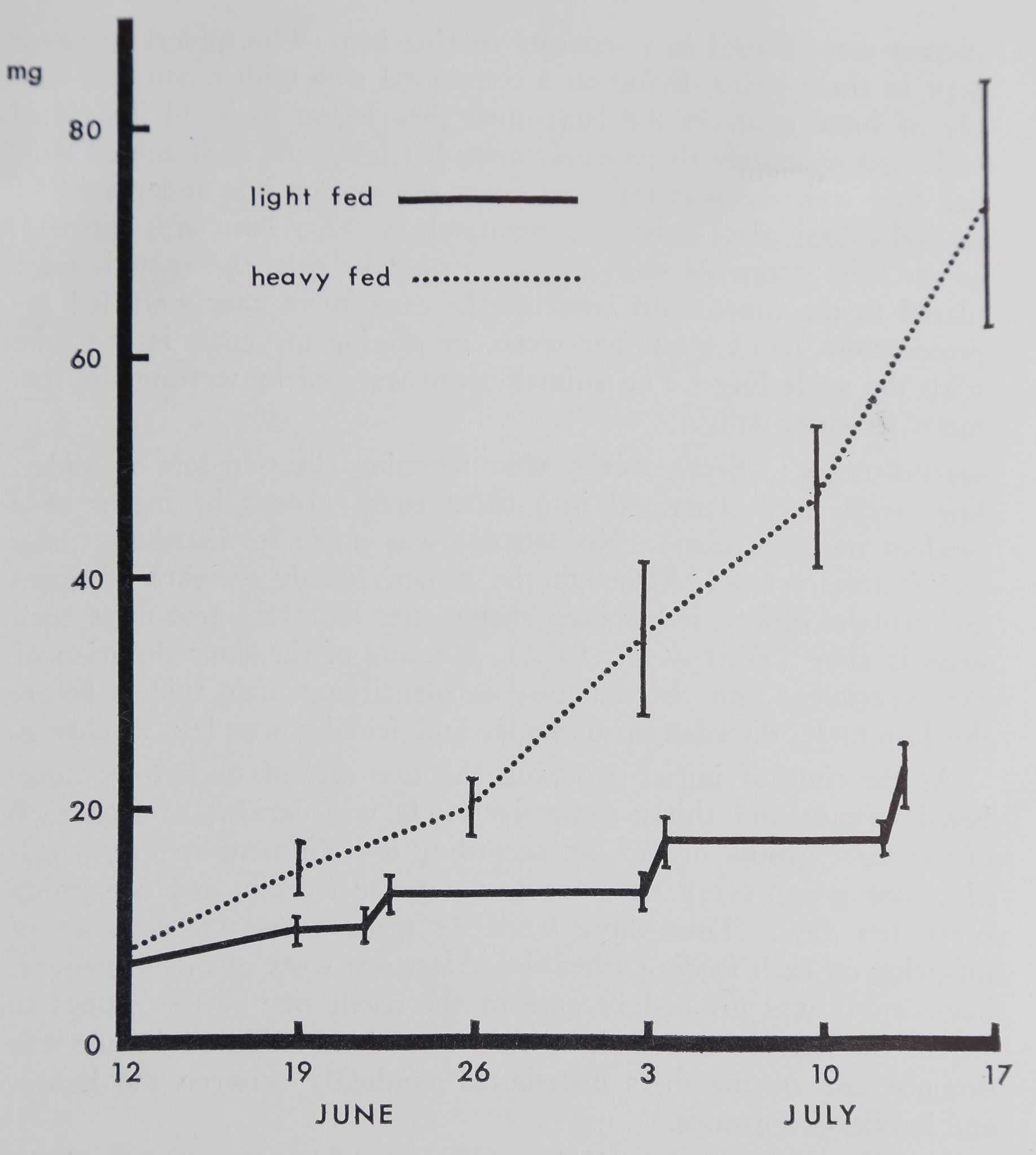
Figure Is Mean weights of 19 light-fed and IS heavy-fed Araneus
diadematus with standard errors (vertical lines). Figures are for both
sets of spiderlings combined. Sharp increases in weight in the light-fed
group are due to the animals being weighed before and after feeding.
Note the increasing difference in weight between the light-fed and heavy-
fed group.
1^73] Benforado & Kistler — Arancus diadematus 93
destroyed daily with the thread left in the cage for the spider to
digest.
feeding: While in the tubes, the spiders were fed by placing a
previously weighed de-winged housefly in the tube daily or every
ten days. Those spiders that would not eat a housefly had three to
seven unweighed gnats placed in their tube. By visual inspection the
following day it was determined whether the fly had been eaten. The
remains of the eaten flies were then weighed to obtain an approxi-
mation of the amount eaten by each spider. The spiders were
watered by wetting the cotton every other day.
After being placed in the cages, if the spider had a web, feeding
was by means of placing the housefly in the web; if there was no
web, we attempted to induce the animal to eat by placing the fly
in front of its mouth. The heavy-fed spiders were offered at least
one fly per day and more, if they would accept it. The light-fed
group was fed one fly once every ten days. If on the day of feeding
of the light-fed group a spider would not eat, a note was made and
the attempt repeated until successful. All spiders were watered on
Mondays, Wednesdays and Fridays by spraying a small amount of
water in each cage.
molts: From the onset of the experiment molts were recorded by
date of the molt to give an indication of the maturation of the
animal.
Results
feeding and weight increase : At the end of a period of five
weeks the two feeding schedules resulted in two significantly differ-
ent weight groups. This development is shown in Figure 1, which
illustrates the increasing difference in weight between the two groups.
At the onset of differential feeding the mean weights of the two
groups were alike, however, a T-test between the mean weights at
the end of the experiment is significant at the 0.1% level.
An analysis of covariance was performed on the data. Because
the original data was non-homogeneous, a transformation [log (x +
10)] was made (Winer, 1962). The initial observation was used
as a covariate in the analysis of covariance. Because the analysis
of covariance indicated no significant difference in the behavior
(growth) of the two families, all figures are for both families com-
bined. For the heavy-fed group the mean weight changed from 7.93
mg zh 1.04 on June 12 to 74-28 mg ± 10.93 on Ji-ily 17. The
mean weight of the light-fed group changed from 6.40 mg rir 0.98
Psyche
[March-J une
94
on June 12 to 17.91 mg ± 2.56 on July 13; there was a significant
interaction between time and feeding schedule below the 1% level.
feeding and maturation: If the number of molts over time is
taken as an indication of speed of maturation, then a relationship
between feeding and rate of maturation can be seen. During the
period of differential feeding the number of molts of the heavy-fed
and light-fed groups differed significantly at the 5% level. The
heavy-fed group had a mean number of 3.0 molts while the light-fed
group had a mean number of 2.3 molts. These results are in agree-
ment with the findings reported by Deevey (1949) with Latrodec-
tus mactans (Fabricius) and indicate that in the laboratory with
only food quantity as a variable, a relationship exists between the
rate of weight increase and the rate of maturation.
initial Weight and rate of growth: From the beginning of
the experiment we noted a wide variation of weights of the individ-
ual animals. At the onset of differencial feeding individual weights
ranged from 1.1 mg to 16.2 mg. In both the light-fed and heavy-
fed groups there existed a positive correlation between initial weight
and final weight. For the light-fed group r — 0.7713 and for the
TABLE I
Measurement Light-fed Early Late Heavy-fed Early Late
Mean wt. (mg) of spiders 12.52 22.30 20.14 59-34
Spiral area (cm2) 118.92 II9-5I OO 00 d\ h-1 H-l 138.32
Center area (mm2) 711.00 877.53 92O.3O I424.3O
Thread length (m) 7-35 7-47 7.56 8.47
Mesh width (mm2) 20.16 22.34 21.79 27.48
Angle regularity 4.25 4.16 5.52 4.62
# of oversized angles 1.67 1.87 2.50 1.80
Relative deviation of spiral turns (South) 0.34 0-33 O.4I 0.35
Selected measurements of webs built by a group of light-fed and heavy-fed
spiders. Because not all spiders built on the same day, early and late webs
of both groups were chosen from two five day periods two weeks apart.
Measurements are divided into those which measure size (above the broken
line) and those which measure regularity. Note the difference between the
light-fed and heavy-fed animals in measures of web size at the late date.
While the heavy-fed group increased in all size measures (see Fig. 2), no
web regularity measures changed. For an explanation of web measure-
ments see Witt et al., A Spider’s Web.
Benforado & Kistler — Araneus diadernatus
95
1973
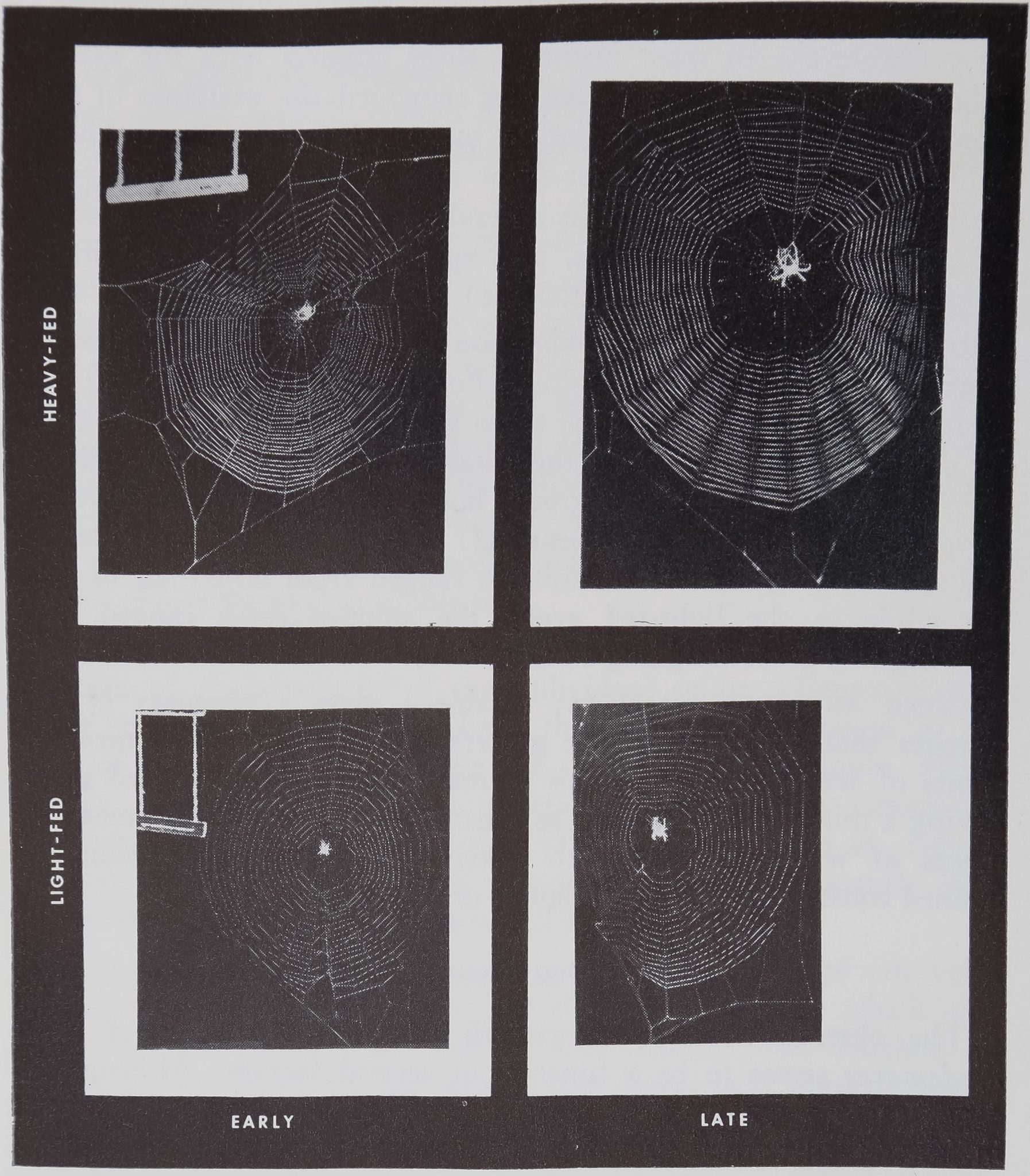
Figure 2. Selected web samples from two spiders: one heavily-fed spi-
der and one light-fed spider. Webs are from the periods measured in
Table 1 and are all reproduced to the same scale. Note that while both
the heavy-fed and light-fed animals began with webs of similar size, after
two weeks of differential feeding the large, heavy-fed spiders’ webs had
increased in size while the webs of the small, light-fed spiders remained
the same size.
Psyche
[ March”} une
96
heavy-fed group r = 0.9m; both correlations are significant at
the 0.1% level. In most instances those animals with the extreme
weights at the beginning of feeding remained the extremes in their
group. Reasons for the variation in initial weight are unknown.
Different rates of growth for light and heavy hatchlings have re-
cently been shown to occur in several species of spiders, apparently
independent of food available, and appear correlated with different
lengths of life (Reed and Witt, 1972).
amount eaten: An approximation of the amount eaten was ob-
tained for a three week period. For nine heavy-fed animals the
mean amount eaten during the three week period was 115.8 mg and
for fifteen light-fed animals the mean amount for the same period
was 35.0 mg. Within each group, however, there was an enormous
variation in the amount consumed : in the heavy-fed group the
amount eaten by individual animals ranged from 209.0 mg to 42.6
mg while in the light-fed group the amount eaten ranged from
49.6 mg to 3 gnats weighing 18 mg.
feeding and web changes: Table I gives a summary of web
changes that accompanied the growth of the animals. In measure-
ments of web size both groups increased, with the heavy-fed group
having a much larger increase as illustrated in Figure 2. In measure-
ments of web regularity both heavy-fed and light-fed groups re-
mained constant, as shown in figures of Table 1.
Discussion
The observed differential growth and development in Araneus
diadematus seems to be a function of several factors. Although an
exposure to a greater than normal supply of food generally results
in faster than normal growth and development, even within a group
exposed to the same food supply there seems to be a great variation
in growth rates. Evidence of these differences is expressed in the
increasing standard errors in Figure 1, and seems to be dependent
upon individual factors in the animals rather than environmental
variations. Large differences in the amount eaten by individual
animals in the laboratory existed and presumably exist in nature.
These differences seem to correspond to differences in the rate of
growth in agreement with the findings of Turnbull in other species
of spiders (Turnbull, i960, 1965). However, whether these dif-
ferences in the amount of food eaten are due to differences in pro-
ficiency in prey-catching or to differences in appetite or some other
factor in the animal is not clarified by our findings.
1973] Benforado & Kistler — Araneus diadematus 97
Another important factor influencing differential growth is the
initial weight of the animal. Variations in initial weights within a
family are generally retained during the course of development. Al-
though several possible reasons for different initial weights within a
family have been given by others, the authors are reluctant to offer
any explanations.
In an orb web weaving spider such as Araneus diadematus the
amount of food available to the animal is roughly equivalent to the
number of prey which become entrapped in the web. The number
of prey entrapped in the web is in turn determined by a number of
variables such as web-site, size and fine structure of the web, and
frequency of web building. Thus, it can be seen that the interaction
of the variables resulting in differential size and growth is complex
and can be divided into those factors which influence the amount of
food available to the spider and those factors which influence the
spider’s use of the food available to it.
Repeated attempts have been made to explain web characteristics
in terms of characteristics of the individual spider (Peters, 1936).
More common, however, has been the notation of changes in the
form of the orb web during the life of the spider (Tilquin, 1942;
Savory, 1952) and the attempt to relate these changes to changes in
the animal (Witt and Baum, i960; Witt, 1963; Reed et al., 1969).
Because influencing factors vary concurrently, it is frequently diffi-
cult to assess the causes of changes in the form of the web.
In our experiment we attempted to isolate the effect of one vari-
able (weight) while minimizing the effect of a variable which nor-
mally changes concurrently (time). All animals used hatched on
the same date, however, one group (the heavy-fed) gained consid-
erable weight over the period measured. The web changes accom-
panying these weight increases are summarized in Table 1. Be-
cause all of the animals were hatched on the same date, we conclude
that increases in web size are due to differences in size of the ani-
mals resulting from differential feeding rather than differences in
age. If appetite were a factor influencing web size, it would appear
that the hungrier, light-fed animals would build a larger web in
an attempt to catch more food; however, this is not the case.
The relationship between food and the web of a spider is a deli-
cate one. Without food, the spider’s web-building ability diminishes,
but without a web there is no food (Peakall, 1968). Thus, like a
businessman, the spider faces the law of diminishing returns. It
appears that the hungry spider chooses to conserve its resources
Psyche
[March-J une
98
rather than gamble on a larger web trapping more food. Early food
deprivation experiments (Witt, 1963) show that the spider con-
tinues to build the same size web when deprived of food, but with
less thread until finally a decreasing in web size occurs. Because
our hungry (light-fed) animals were kept on a diet closer to a main-
tenance level than a deprivation level, we observed no decreases in
web dimensions.
Feeding conditions in a natural environment vary more than those
imposed in a controlled laboratory. Yet the spider is able to survive
in these naturally diverse conditions because of its adaptability. In
situations where there is little food available, the spider is able to
survive by growing at a slow rate and maintaining the same size
web. Where food is abundant, the spider takes advantage of the
situation, growing at a fast rate and increasing the size of its web.
The spider has developed a method for coping with a wide range
of feeding conditions. By varying its body and web growth, the
spider can survive under the diverse conditions imposed by nature,
thus minimizing the necessity of seeking new food supplies and re-
locating the web. Our findings provide new insight into the spider
as an example of an animal that adapts itself successfully to its en-
vironment.
Summary
Spiders from two cocoons of Araneus diadematus were exposed to
five weeks of two different feeding schedules: one group was of-
fered large amounts (one housefly per day) of food, the other group
scarce (one fly every ten days) amounts. Although both groups in-
creased in weight, weight gains of the heavy-fed group were signifi-
cantly greater than those of the light-fed group, regardless of cocoon
origin. Within each group there was a wide variation in the growth
of individual animals, indicating the presence of factors other than
food supply; i.e. animals with extreme weights within a group at
the onset remained the extremes.
In conjunction with increases in weight, over the three week period
of observation, webs of the heavy-fed spiders showed an increase in
size but not in regularity and shape in comparison to webs of the
smaller, light-fed animals of the same age which did not change.
Such data suggest an increased chance of survival of the species
through variations in rate of growth and maturation dependent on
environmental factors.
Benforado £sf Kistler — Araneus diadematus
99
1973]
Acknowledgements
This work was carried out in the laboratories of the North Caro-
lina Department of Mental Health and was supported by Grant
Number GB 25274 from the National Science Foundation to Peter
N. Witt. The authors gratefully acknowledge the assistance of Dr.
Peter N. Witt during all stages and the assistance of Mrs. Mabel B.
Scarboro during the period of laboratory work.
References Cited
Bristowe, W. S.
1958. The World of Spiders. Collins, London.
Comstock, J. H.
1940. The Spider Book, Revised and edited by W. J. Gertsch. Com-
stock, Ithaca, N.Y.
Deevey, G. B.
1949. The development history of Latrodectus mactans (Fabr.) at dif-
ferent rates of feeding. Amer. Midland Naturalist. 42: 189-218.
Levi, H. W.
1971. The Diadematus group of the orb-weaver genus Araneus north
of Mexico (Araneae: Araneidae). Bull. Mus. Comp. Zool., 141:
131-179.
McCook, H. C.
1890. American Spiders and Their Spinningwork. Vol. 2, Published
by the author, Philadelphia.
Peakall, D. B.
1968. The spider’s dilemma. New Scientist, pp. 28^29.
Peters, H. M.
1936. Studien am Netz der Kreuzspinne (Aranea diadema.) 1. Die
Grundstruktur des Netzes und Beziehungen zum Bauplan des
Spinnenkôrpers. Z. Morphol. Ôkol Tjere, 32: 613-649.
Reed, C. F. and P. N. Witt
1972. Growth rate and longevity in two species of orb-weavers. Bull.
Brit. Arach. Soc., 2: 111-112.
Reed, C. F., P. N. Witt and R. L. Jones
1965. The measuring function of the first legs of Araneus diadematus
Cl. Behavior, 25 : 98-119.
Reed, C. F., P. N. Witt and M. B. Scarboro
1969. The orb web during the life of Argiope aurantia (Lucas).
Develop. Psychobiology, 2: 120-129.
Savory, T. H.
1928. The Biology of Spiders. Sidgwick and Jackson, London.
1952. The Spider’s Web. Frederick Warne and Co., London and N.Y.
Tilquin, André
1942. La Toile Géométrique des Araignées. Presses Universitaires de
France, Paris.
Turnbull, A. L.
1960. Quantitative studies of the food of Linyphia triangularis (Clerck)
(Araneae: Linyphiidae). Canad. Ent. 94: 1233-1249.
100
Psyche
[March-June
Turnbull, A. L.
1965. Effects of prey abundance on the development of the spider
Agelenopsis potteri (Blackwell) (Araneae: Agelenidae). Canad.
Ent. 97: 141-147.
Winer, B. J.
1962. Statistical Principles in Experimental Design. McGraw-Hill,
N.Y. pp. 606-615,
Witt, P. N.
1963. Environment in relation to behavior of spiders. Arch. Environ.
Health, 7: 4-12.
1971, Instructions for working with web-building spiders in the lab-
oratory. BioScience, 21: 23-25.
Witt, P. N. and Ricarda Baum
1960. Changes in orb webs of spiders during growth. Behavior, 16:
309-318.
Witt, P. N., J. O. Rawlings and C. F, Reed
1972. Ontogeny of web-building behavior in two orb-weaving spiders.
Am. Zoologist, 12: 445-454.
Witt, P. N,, C. F. Reed and D. B. Peakall
1968. A Spider’s Web. Springer Verlag, Berlin.


