Warning: Trying to access array offset on value of type null in /home/elisew5/public_html/drpeterwitt.com/wp-content/plugins/contextual-related-posts/includes/content.php on line 49
Images Collection
View this article in Search Friendly Plain Text
NOTE: This plain text article interpretation has been digitally created by OCR software to estimate the article text, to help both users and search engines find relevant article content. To read the actual article text, view or download the PDF above.
STUDIES ON VERATRUM ALKALOIDS. XXV. VERATRINE RESPONSE AND ANTIVERATRINIC ACTION IN FROG SARTORIUS MUSCLE
PETER N. WITT and CHARLES R. SWAINE Department of Pharmacology, Harvard Medical School, Boston, Massachusetts
Reprinted from The Journal op Pharmacology and Experimental Therapeutics Vol. 120, No. 1, May, 1957 Printed in U.S.A.
Reprinted from Tub Journal op Pharmacology and Experimental Therapeutics Vol. 120, No. 1, May, 1057 Printed in U.S.A.
STUDIES ON VERATRUM ALKALOIDS. XXV. VERATRINE RESPONSE AND ANTIVERATRINIC ACTION IN FROG SARTORIUS MUSCLE1
PETER N. WITT2–3 and CHARLES R. SWAINE Department of Pharmacology, Harvard Medical School, Boston, Massachusetts
Received for publication December 11, 1956
The veratrine response of the amphibian muscle is caused by the ester alkaloids, veratridine and cevadine, from Schoenocaulon officinale. Since these ester alkaloids are now available it seems advantageous to abandon the use of the ill-defined mixture “veratrine” and employ one of the ester alkaloids. In previous work from this department veratridine was used and has been found suitable to reproduce the veratrine response (Krayer and George, 1951; Arora and Krayer, 1952; Arora, 1953; Fuentes, 1957).
In the characteristic myogram of the frog sartorius muscle treated with veratridine the initial twitch-like contraction resulting from a single electrical stimulus is followed by a more or less pronounced relaxation; this is followed again by a slow secondary rise and fall of tension. Sometimes with high concentrations of the alkaloid the secondary rise is superimposed upon the initial twitch without any intervening relaxation. It is the secondary rise and fall of tension which is referred to in this paper as the veratrine response (VR).
The secondary alkamines among the veratrum alkaloids, such as veratramine from Veratrum album or Veratrum viride, do not possess veratrinic action, rather they abolish the VR and restore the normal twitch. In this regard they are similar to quinine and quinidine (Krayer and George, 1951). The term “antiveratrinic action” was introduced by Krayer to characterize substances which, in appropriate dosage range, are capable of antagonizing, in the presence of veratridine (or a similarly acting substance), the secondary rise of tension without reducing the initial twitch.
An aid in the investigation of the VR was the application of the intracellular microelectrode technique as described by Nastuk and Hodgkin (1950). This method enabled us to study the fast electrical changes of the veratrinized muscle in the single cell. The combination of this technique with simultaneous recording of tension of a small group of fibers should clarify the question whether there exists a close connection between the electrical changes in the cell membrane (seen as repetitive discharges and negative afterpotential) and the characteristic VR.
Methods. Prog sartorius muscle was prepared so that no fiber was impaired. For most of the experiments reported here the size of the muscle was reduced by cutting a certain
1 This work was supported in part by funds from the Eugene Higgins Trust and in part by a research grant (H-2205) from the U. S. Public Health Service.
2 Rockefeller Fellow.
3 Present address: Department of Pharmacology, State University of New York, Upstate Medical Center, Syracuse, N. Y.
WITT AND SWAINE
()4
number of fibers in the middle, pulling the cut ends back to their respective insertions at both ends, and eliminating them carefully. The connective tissue which surrounds the muscle was removed as completely as possible. Nearly all of the experiments were made with approximately 100 intact fibers. The number of intact fibers could be roughly estimated by the tension developed in response to a supramaximal stimulus.
The buffered Ringer solution used was prepared according to Nastuk and Hodgkin’s “Normal Ringer” (1950). Pure oxygen, air or the gases indicated under the respective experiments were bubbled through the bathing solution during the experiments.
The dissected muscle was fixed with its proximal end near one side of the bottom of the vessel containing the Ringer solution in an angle of 45° to the horizontal. The distal one-sixth of the muscle was not immersed, but was kept wet with Ringer solution, and was connected by a glass rod to a strain gauge. The stimulating platinum electrodes were fixed to the distal end. By adjusting initial tension it was possible to determine that tension which produced a maximal twitch after a supramaximal stimulus. Smaller variations in initial tension showed no detectable change in the sensitivity to veratridine and the form of the VR.
Tension was recorded by means of a Grass strain gauge, amplifier, demodulator and inkwriter (especially adapted for this purpose). The inertia of this transmitting system was found low enough to give a full sized excursion with a 1 msec, sine wave. The calibration was not fully linear as can be seen from figures 1 and 2. The amplification which remained the same throughout the experiment could be regulated for each experiment according to the tension which the muscle developed. The speed of the recording paper could be increased up to 60 mm./sec., allowing tension measurements every 0.0166 sec. This recording device did not enable us to measure the latency period between the stimulus and the development of tension. It is assumed that for figures 1 and 2 the latency is of the order determined by Sandow (1952).
Supramaximal stimuli of 0.1 msec, duration were delivered from a Grass stimulator, model 4. As intensity was never increased, our results include only experiments with muscles in which the excitability was not markedly decreased. As a control, each stimulus transmitted to the muscle was recorded simultaneously on the inkwriter. Stimulus duration was kept at 0.1 msec, to minimize the stimulus artifact. As the recording microelectrode was placed into a distant part of the muscle, only conducted impulses were measured.
Microcapillaries prepared as described by Ling and Gerard (1949), with a tip diameter of two miera or smaller and filled with saturated KC1 solution, were lowered into a fiber by means of a de Ponbrune micromanipulator (de Fonbrune, 1949) under optical control through a stereoscopic microscope. The microelectrode was connected through a 25 mm. long piece of polyethylene tubing filled with KC1 and an unpolarizable silver-silver chloride electrode to the grid of a cathode follower. The cathode follower with the tube 5800 from the Victoreen Instrument Company was set up as described by Nastuk and Hodgkin (1950). Its input valve had a grid current of 6.8 X 10“11 amp. The second electrode was a saturated KCl-agar filled glass tube which dipped into the Ringer solution. It was connected through another silver-silver chloride electrode. The voltage was called zero when both electrodes dipped into the Ringer bath.. The voltage was amplified through a two channel DC amplifier4 * and connected to the plates of a Dumont flatfaced cathode ray oscilloscope. The movements of the ray were recorded with a Grass Kymograph Camera6. The film speed was 10 cm./sec. and figures 1 and 2 show the original records in low magnification. Each microelectrode was checked for its resistance and the whole apparatus for its ability to transmit the expected changes of voltage undistorted in time. When a voltage step was introduced in series with the microelectrode the new potential was reached with a time constant of 0.1 msec.
4 Lent to us by the Department of Pharmacology, University of Bern, Bern, Switzer
land, through the kindness of Dr. W. Wilbrandt.
6 We wish to express our gratitude to Mr. A. M. Grass of the Grass Instrument Co., Quincy, Mass, for kindly lending us the oscilloscope and the camera.
VERATRINE RESPONSE IN FROG MUSCLE
65

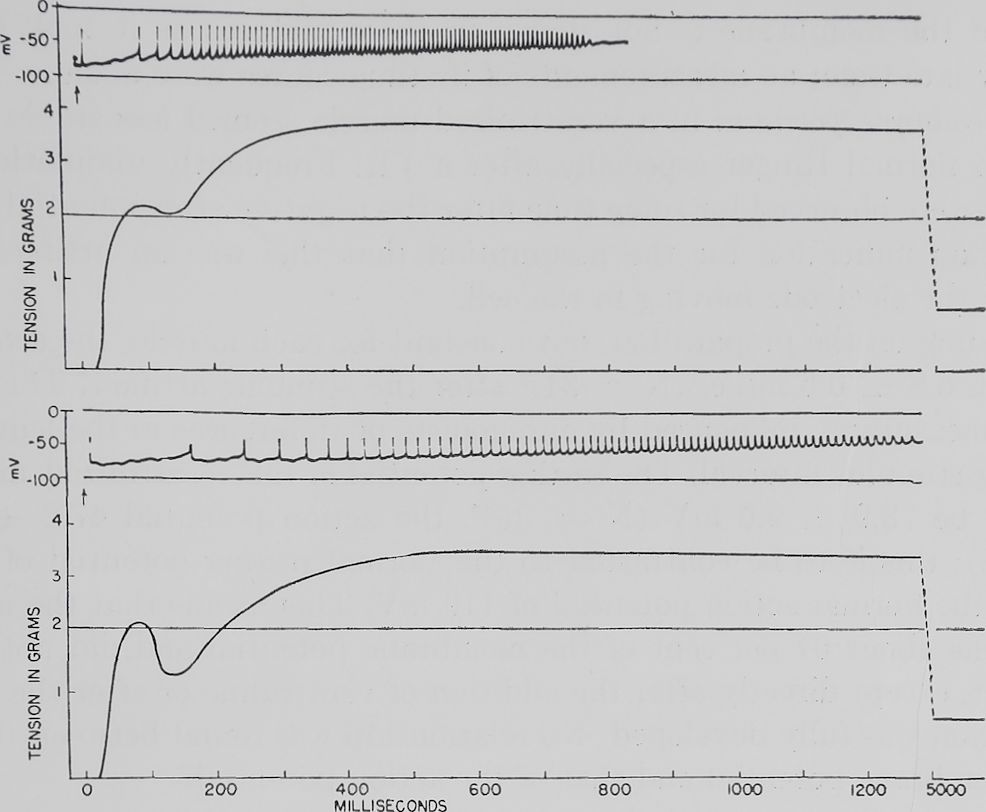
Fig. 1. Effects of veratridine upon electrical and mechanical events in striated muscle.
Rana pipiens, male. Fiber bundle of sartorius muscle. Two records of veratrine responses are shown: both electrograms are from the same single fiber; the corresponding myograms are from the whole fiber bundle. The upper response was obtained by a stimulus (at the arrow) given to the fiber bundle after a two-minute rest period. The stimulus (at the arrow) of the lower response was given one minute later.
The veratridine used was the free base prepared from veratrine6 by Deliwala and Kup-chan. It had a rotation of [a] Is + 6° (c = 2.00 in ethanol). Stock solutions containing 1 mgm./ml. were prepared by dissolving the alkaloid in distilled water with an equimolar amount of hydrochloric acid. The veratramine, which was generously supplied by O. Wintersteiner of E. It. Squibb & Sons, was dissolved in the same way as veratridine to produce a stock solution of 10 mgm./ml. In most experiments appropriate amounts of the stock solution were added to the bath to obtain the concentration desired. In some experiments the whole bathing solution was replaced.
In the beginning of the experiments the microelectrode was often broken or pulled out of the cell by the movement of the muscle during a VR. Later on this difficulty was largely overcome by choosing an electrode with a thin, elastic tip and by reducing the number of muscle fibers which led to a reduction in pull during the twitch. If after a VR the membrane potential returned to the old value and the cell had shown action potentials, the photograph was used for evaluation. However, from time to time strange forms of potential changes appeared and it could not be decided whether they were artifacts due to our recording method or real potential changes in the cell membrane. These photographic records were not included in our evaluation.
Results. The activity of a muscle fiber during a VR. Figures 1 and 2 represent typical tracings of the photographs obtained during a VR. Shortly after the stimulus was given (represented by the stimulus artifact above the arrow) the action potential rose sharply, fell approximately to the preceding membrane potential, and was followed by a number of repetitive discharges which arose from a slowly decreasing membrane potential, the negative afterpotential. The repolar-
8 Generously supplied by J. B. Penick & Co., New York, N. Y.
WITT AND SWAINE
66
ization of the membrane is not shown in these pictures as it took place at a relatively late timej no measurements of its time course were made.
The membrane ‘potential in a veratrinized muscle seemed less stable than in a muscle in normal Ringer especially after a VR. Frequently undulations of the potential were observed for some time after the negative afterpotential was over. There is no indication for the assumption that this was an artifact brought about by the electrode moving in the cell.
Depending on the preparation, but constant for each muscle, the action potential began 6.8 dr 0.6 msec. (N = 34)7 after the stimulus artifact. This distance was not measurably influenced by antagonists or differences in the length of the preceding stimulus interval. The resting potential in the veratrinized muscle was found to be 73.9 ± 9.6 mV (N = 35)8, the action potential 47.2 ± 1.8 mV (N = 35); this is to be contrasted to the normal resting potential of 86 to 88 mV and the normal action potential of 119 mV. This means that the action potential was about 64 per cent of the membrane potential and did not show an overshoot, except directly after the addition of veratridine or after the antivera-trinic action was fully developed. No relationship was found between the height of the membrane potential and that of the action potential.
While the action potential showed a rather constant time distance from the stimulus, the repetitive discharges began after a widely varying time interval (see example in figure 1). The most frequently used stimulus interval of two minutes showed an average time between the stimulus artifact and the first discharge of 109 ± 21 msec. (N = 43). The repetitive spontaneous electrical discharges following a four-minute stimulus interval showed an average time of 38.6 ± 32.9 msec. (N = 3); and the discharges following the one minute interval came after an average time of 128.8 ± 9.8 msec. (N = 13). The three values are not significantly different which probably can be attributed to the relatively small number of observations.
The time interval between the individual discharges in one VR was—with very few exceptions—regularly decreasing, sometimes constant, seldom irregular. The average distance between the discharges in one VR was measured as the distance between two potentials which lay in the middle of the total number. This seemed again dependent on the preceding stimulus interval, being 19.5 zb 5.0 msec. (N = 10) for the one-minute interval, 13.4 ±4.1 msec. (N = 42) for the two-minute stimulus interval, and 7.2 ± 0.7 msec. (N = 5) for the four-minute interval. The results varied widely from fiber to fiber but records from the same fiber consistently showed a shorter average distance after a longer resting period. An example is shown in figure 1.
The average height of the first action potential of a group of repetitive discharges measured from its take-off level was 39.0 ± 2.3 mV (N = 74)9, being
7 In this paper the first figure is the mean: the figure following ± is the standard error of the mean; N is the number of observations; the concentrations are in w/v.
8 For evaluation we used records of all fibers that showed action potentials and in which the membrane potential had the same height before and after a VR.
9 This figure includes some measurements of veratrine responses soon after the anti-veratrinic substance was given since the height was not measurably influenced at first.
VERATRINE RESPONSE IN FROG MUSCLE
67

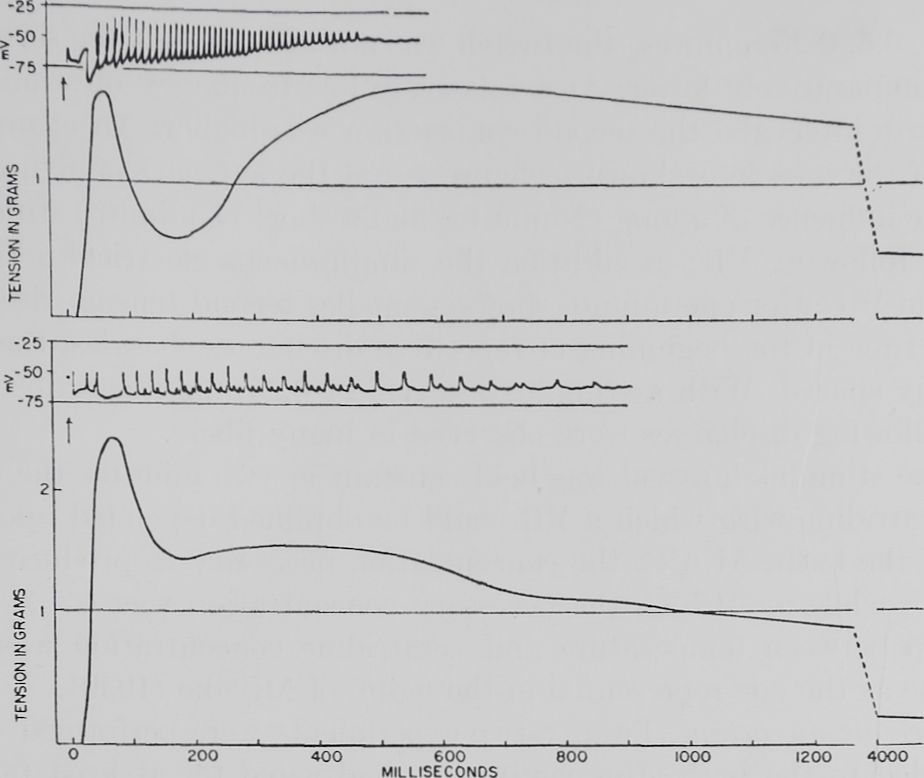
Fig. 2. Effect of veratramine on the electrical and mechanical events of the veratrine response.
Rana pipiens, male. Fiber bundle of sartorius muscle. The electrograms are from two different fibers in the same fiber bundle; the corresponding myograms are from the same whole fiber bundle. Both responses were obtained after a two-minute rest period. The veratrine response shown in the lower tracing was obtained 13 minutes after veratramine had been added in a concentration of 10“6.
nearly always lower than the preceding action potential. The following discharges in one VR always showed a decrease in height, mostly regularly decreasing. Discharges increasing in height were never observed by us. That the discharges decreased in height was partly due to the decrease in membrane potential —which means a decreasing level of the take-off voltage—and partly due to a lower amplitude of the discharge.
The total number of discharges varied widely and up to 166 were recorded by us. There was some difficulty in obtaining exact figures of the total number as during many veratrine responses the height of the discharges continuously decreased until only a damped oscillation of the membrane potential was discernible, and it was sometimes impossible to decide which discharge should be considered the last one. In other veratrine responses the discharges ended abruptly. The average figure for the total number of discharges in our experiments was 33.4 ± 3.2 (N = 64). The number showed no relationship to the preceding stimulus interval. In each muscle some fibers showed no discharges at all, but our technique did not enable us to decide how far these fibers took part in the VR.
The effect of stimulus frequency, temperature and drug concentration on the VR. The influence of the resting period preceding a stimulus on the VR was investigated in 59 experiments. As is well known, a change in the frequency of stimulation is followed by a change in the VR: by applying the stimulus with shorter
WITT AND 8WAINE
08
interval (1, 0.5, 0.25 minutes) the twitch was not affected but the second tension rise was comparatively lower. At a relatively low frequency of stimulation (4-, 8-, 16-minute intervals) the second contraction was higher. An example of this behavior of the muscle is shown in figure 1 and the left part of figure 3. Figure 1 shows the influence of a long (2-minute) and a short (1-minute) stimulus interval on the following VR; in addition the simultaneous electrical activity is recorded. The VR after one minute shows a smaller second tension rise as well as a delay in time in the beginning of repetitive discharges; besides, the latter are more widely spaced. With a stimulus interval of 0.25 minute, action potentials without following discharges were observed in many fibers.
When the stimulus interval was held constant at two minutes the concentration of veratridine with which a VR could be obtained depended upon the temperature of the bath. At 6°C. the concentration necessary to produce a VR was 1.3 X 10“6, while at 31.5°C. the necessary concentration was 1.3 X 10-8. The relationship between temperature and veratridine concentration was found to be the same as the one represented in the figure of Miyake (1939).
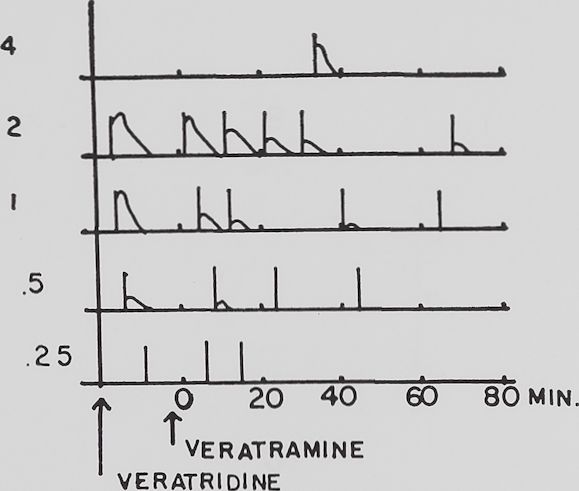
The antiveratrinic action. Twenty-two experiments were performed with verat-ramine added to the bath after veratridine had acted for at least ten minutes at a time when the muscle showed its characteristic VR. The substance was added to the bath in an amount to obtain an end concentration of 10“5. Stimuli were applied to the muscle regularly every two minutes. After an average time of 26 minutes in a whole sartorius muscle and an average time of 8.3 minutes in a 100 fiber preparation, the VR reached only one-half of its initial height. The difference in time between the whole muscle and the 100 fiber preparation is significant below the 5 per cent probability level with t = 2.87. The influence of different stimulus frequencies, as described in the last paragraph, could be obtained at any time during the veratramine antagonism. This means a second tension rise in a VR reduced by veratramine action could be brought back again to its full initial height after waiting 4 or 8 minutes between stimuli instead of 2. The addition of veratramine has an effect which can be described as equal to that of shortening the stimulus interval. This is illustrated in figure 3.
The antagonized VR, the short muscle twitch without a second tension rise, often looked electrically like an action potential rising from a higher membrane potential. A few records of the electrical behavior of the cell were obtained during the transient period in which the second tension rise was already smaller but not yet totally abolished. At this time only a small number of fibers showed repetitive discharges and these were changed in shape as shown in figure 2. The discharges were irregular in height and some spikes showed an abnormally slow time course.
After having obtained an antagonized VR with veratramine the concentration of neither veratridine nor veratramine could be increased without suppression of the activity of the muscle. There was no indication for competition between the two substances. While the concentration of veratridine necessary to elicit a full VR changed with the temperature, as soon as a VR was obtained at a certain temperature, it could always be antagonized with veratramine at 10“B. Only the
VERATRINE RESPONSE IN FROG MUSCLE
69
effective veratridine concentration was temperature-dependent, not the antiver-atrinic veratramine concentration.
Strophanthin K (ampules Strophosid Sandoz) was used in 9 experiments. The final concentration in Ringer was between 2 X 10~6 and 3 X 10“6. The Strophanthin had been added 14 minutes before the veratridine was given and a stimulus was given every two minutes, as described by Arora (1953). The results were similar to those obtained with veratramine. The influence of muscle diameter, temperature and the electrical behavior of the cells were not investigated.
Quinine and quinidine (12 experiments) showed an antiveratrinic action in a concentration of 10-6, and the effect had» the same characteristics described in the veratramine and Strophanthin experiments. After the quinine had begun to act on the VR, a large action potential with overshoot and without repetitive discharges could be recorded. However, in two photographs taken during the
MIN

Fig. 3. Influence of rate of stimulation and veratramine on the veratrine response.
Rana pipiens, male. Sartorius muscle. Ordinate: tension. The figures refer to the stimulus interval preceding each veratrine response. Abscissa: time in minutes after the addition of veratramine. The veratrine responses are traced from the original records.
transient decline of the VR the electrical activity of the cell could be shown to be the same as that observed after veratramine shown in figure 2.
Iodoacetic acid (18 experiments) was shown to act on the VR in a manner different from that of veratramine, Strophanthin and quinine. After having-added iodoacetate 10~4 to the Ringer-veratridine bath, the VR remained unchanged for several minutes then both—the first quick rise of tension and the VR—gradually declined in height. The rate of the tension decrease was dependent on the initial height of the VR and was not always equal for both parts; but in no case was only the VR affected. According to our definition therefore iodoacetic acid has no antiveratrinic activity.
The influence of dinitrophenol (7 experiments in concentrations between 10~8 and 10~6), sodium cyanide (9 experiments at 10~4) and nitrogen (3 experiments) on the VR followed the pattern described for iodoacetate. The time constant of the effect varied with different concentrations and different substances. By vary-
70
WITT AND SWAINE
ing the concentration and stimulus frequency no indication could be found that any one of these substances had antiveratrinic activity. A representative experiment is plotted in figure 4B.
The influence of sodium excess in the Ringer solution (120 per cent to 135 per cent as compared to normal Ringer 100 per cent) on the VR was investigated

TIME N MINUTES
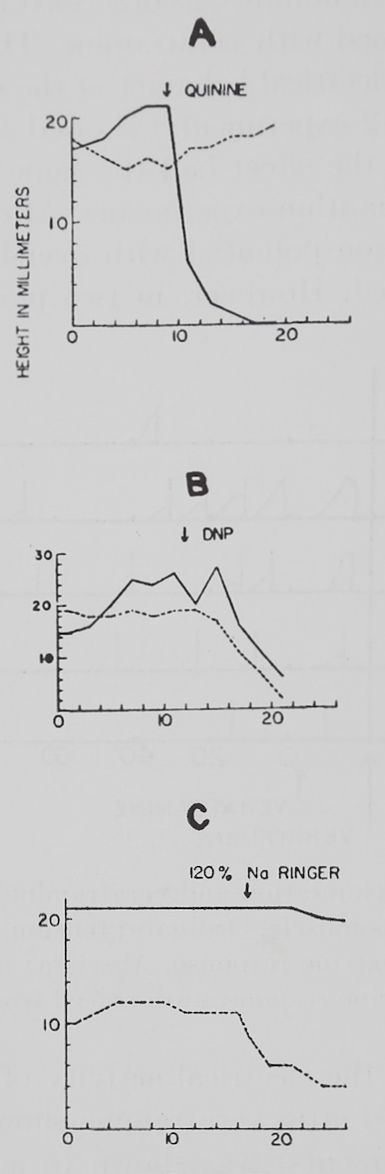
Fig. 4. Three ways in which substances influence the veratrine response.
Rana pipiens, male. Sartorius muscle. Ordinate: tension as measured in mm. on the original record. Abscissa: experimental time. Solid line: height of second tension rise. Broken line: height of twitch. A: In A, quinine abolished the second tension rise while the twitch remained constant. B: In B, dinitrophenol caused both the twitch and the second tension rise to decrease simultaneously. C: In C, excess sodium caused the twitch to decrease while the second tension rise remained relatively constant.
in 5 experiments. The pattern of the influence of sodium excess on the VR was found different from both patterns described earlier. Figure 4C shows a representative experiment. It can be seen that the known decrease of twitch height in Ringer with excess sodium does not coincide with a decrease of the VR. On the other hand, in some experiments the VR increased simultaneously with the
VERATRINE RESPONSE IN FROG MUSCLE
71
decrease in twitch height. This effect lasted only 15 minutes; and no systematic studies were made over a long period of time. Only in one experiment electrical records were obtained from the muscle fibers. The variation of the data obtained from different fibers was too great to make valid statements about the height of the membrane potential, the action potential, or the repetitive discharges. However, the increase of the VR under sodium excess corresponded electrically to a higher number of repetitive discharges per unit time and a decrease in distance between the stimulus artifact and the first action potential when compared to the control VR in the same muscle.
Decamethonium bromide was added in 4 experiments in which the muscle preparation under veratridine was stimulated alternately through the nerve and directly at the muscle tissue. At a concentration of 5 X 10~6 decamethonium inhibited the effect of indirect stimulation while direct stimulation still produced an unchanged VR. In two of the four experiments, exchanging the bathing solution for normal Ringer resulted in a restoration of the VR when the muscle was stimulated through the nerve. Electrical records taken after the addition of decamethonium 10-4 in one experiment showed that repetitive discharges occurred.
Discussion. In our veratridine experiments the average membrane potential was 73.9 ± 9.6 mV as compared to 88 ± 1 mV found by other authors in the normal frog sartorius muscle (Nastuk and Hodgkin, 1950). We have no measurements of our own with unpoisoned muscles. Straub (1956) found the same effect of veratridine on the resting potential of myelinated nerve fibers of the frog. The membrane potential in each cell showed a stable value to which it returned after a VR. However, a wide variation existed in height of membrane potential from cell to cell, and some abnormally high and low values were observed. There seemed to be no relationship of the height of the resting potential to either the VR or the ability of the cell to produce repetitive discharges. Similar findings in nerve fibers were reported by Guttman (1940), Acheson and Rosenblueth (1941) and Hodler, Staempfli and Tasaki (1950). It may indicate that not the height of the membrane potential but its instability is characteristic for the action of veratridine in low concentrations.
The amplitude of the action potential appeared distinctly decreased under the conditions of our experiments: 47.2 ± 1.8 mV as compared to 119 ± 1 mV (Nastuk and Hodgkin, 1950). This is also confirmed by measurements on the nerve by Graham and Gasser (1931) and Schmitt and Gasser (1933). The height of the action potential seems independent of the increased twitch height under veratridine as was found already by Rosenblueth et cd. (1941). It seems the more probable that the low action potentials which we found are attributable to the veratridine, since development of the full antiveratrinic action sometimes brought back overshooting action potentials on a higher membrane potential.
If the correlation between frequency of repetitive discharges, stimulus interval, and steepness of second tension rise, as shown in figure 1, could be confirmed in further experiments (possibly the two first properties also in nerve fibers) it would mean that immediately after a VR the muscle becomes refractory to a second VR, but not to a single action potential. The longer the interval between
72
WITT AND SWAINE
stimuli the greater the VR. The membrane of the veratrinized muscle behaves like that of the nerve when stimulated with alternating current (Schaefer and Goepfert, 1937). The characteristic of the membrane which determines the frequency of conducted impulses would be dependent on the rest period which the muscle had after the last VR. That the repetitive discharges become smaller and at last cease is also consistent with the findings of Schaefer and Goepfert (1937) on the normal nerve membrane. They describe a slowly developing inhibition of conducted stimuli at high frequency stimulation.
No definite statements about excitation-contraction coupling in the VR can be made on the basis of our experiments since our tension records are not from single fibers. However, the parallelism between the steepness of tension rise and density of repetitive discharges appears striking enough (figure 1). There seem to be no parallel measurements about the phenomenon of dependence of frequency of discharges and the preceding resting period in nerve fibers. If this relationship could be found consistently it would be a further indication that veratridine acts on the excitable mechanisms of the membrane, and the contractile processes only follow the changes in the membrane without dissociation. This statement does not refer to the relaxation of the muscle in the VR, as we did not register the negative afterpotential in its end phase.
If the assumption is correct that veratridine acts on the excitable mechanisms of the cell membrane it should be inactive when applied to the isolated contractile substance. Robb (personal communication) made such experiments with actomyosin threads with a technique described previously (Robb and Mallov, 1953). Robb found that “veratridine in a concentration which affects the whole muscle fiber does not enhance shortening of weighted threads to a degree which is statistically valid.”
The old observation could be confirmed that neither the nerve nor the end-plate are necessary for the VR of skeletal muscle. The result of the decametho-nium experiments seems of interest since this substance blocks neuromuscular transmission at the endplate in a different way from that of curare (Patón and Zaimis, 1952). The depolarization of the membrane in the endplate region by decamethonium obviously did not interfere with the origination of a VR in muscle fiber.
Considering the substances which show no antiveratrinic activity, several metabolic processes become unlikely as being directly affected by varatridine. It is improbable that veratridine interferes primarily with the oxidative processes, because lack of oxygen (experiments in nitrogen atmosphere) and blocking of oxygen transport (sodium cyanide experiments) do not interfere specifically with the VR. Likewise there seems no direct interference with the acetylcholine metabolism (Grupp, 1951). The experiments of Sandow and Kiebel (1952) can be interpreted in the same way if the antiveratrinic action is defined as strictly not affecting the twitch height. The results of the experiments with iodoacetic acid and dinitrophenol would seem to indicate that veratridine affects neither the formation of lactic acid nor the oxidation-phosphorylation coupling in muscle.
The group of antiveratrinic substances is chemically not homogeneous. The
VERATRINE RESPONSE IN FROG MUSCLE
73
cellular mechanism of action of cardiac glycosides as well as of quinine and ve-ratramine is unknown and consequently little can be said concerning the mechanism of their antiveratrinic action. Recent work done with Strophanthin on erythrocytes (Schatzmann, 1953) and frog skeletal muscle (Schatzmann and Witt, 1954) suggests that digitalis glycosides interfere with an active ion transport mechanism in the cell membrane. As Strophanthin is an antagonist of verat-ridine the ion transport mechanism across the cell membrane might be the point of attack of both substances, however, the data indicate that such an antagonism would not be competitive in nature.
The change in the VR after antiveratrinic substances seemed not to be brought about only by a participation of fewer and fewer fibers in the VR. Our single cell records indicated that there existed, at least in many fibers, a transient state in which the VR in each fiber is diminished through a diminished frequency of repetitive discharges from each cell. If the lower parts of figures 1 and 2 are compared there seems to be a certain similarity in the action of a short stimulus interval and antiveratrinic action. In both cases the frequency of repetitive discharges is diminished. The antiveratrinic substances may also increasingly prolong the period in which the muscle is refractory to a VR, as was suspected in the discussion of the result of a small stimulus interval. The action of veratri-dine appears to be coupled to a mechanism which is active in the restoration period after activity. There exists another indication that the veratridine action is coupled to a process in connection with the frequency of muscle activity. Benforado and Witt (1954) observed that the lethal dose of veratridine infused into a heart was dependent on the heart rate; faster beating hearts went into fibrillation at an earlier time, i.e., after the administration of a smaller lethal dose.
An unstable membrane potential, a low action potential, repetitive discharges depending probably in their frequency on the length of the preceding recovery period, the indications for a close parallelism between excitation and contraction, the probable indifference of the contractile substance itself to veratridine, the influence of sodium excess on the VR, and antiveratrinic activity—all these seem to indicate that veratridine affects a mechanism related to the excitation of the cell membrane. This mechanism would be responsible for maintaining the right concentrations of ions on both sides of the membrane and would be responsible for the renormalization of the concentrations after an activity. Consequently, more direct data than electrical records, namely measurements of ionic flux into and out of the cell, should provide the information necessary to elucidate the mechanism of action of veratridine on the muscle cell.
SUMMARY
Simultaneous records were made of the isometric tension developed by about 100 fibers of the frog sartorius muscle and the electrical activity of a single fiber.
Administration of veratridine 10~7 resulted in a state of the muscle where a single stimulus was followed by the characteristic two-peaked tension response
74
WITT AND SWAINE
simultaneous with a series of electrical discharges; these were superimposed on a negative afterpotentiaJ. The second tension rise (VR) and the frequency of firing was higher (7.2 msec, average distance) when the muscle had been allowed to rest for four minutes instead of for only one minute (19.5 msec, average distance). The membrane potential under veratridine was found relatively low (73.9 mV) and unstable, the action potential without an overshoot (47.2 mV).
Addition of veratramine 10-5, Strophanthin 3 X 10~6, quinine or quinidine 10~5 to the veratrinized muscle normalized the situation in that a single stimulus was again followed by a single action potential and twitch. In the transient period when the full antiveratrinic action had not yet developed, unusually slow repetitive discharges were observed together with a small second tension rise.
At lower temperatures (6°C.) higher veratridine concentrations (1.3 X 10“6) were necessary to elicit a VR, but the antiveratrinic action of veratramine was at all temperatures fully developed at 10~5.
Lack of oxygen, addition of sodium cyanide, iodoacetic acid, and dinitrophenol reduced the amplitude of both parts of the mechanical response simultaneously; they had no antiveratrinic action.
Sodium excess in the Ringer bath diminished the twitch before it influenced the VR.
Like curare, decamethonium did not affect the VR and repetitive discharges in a concentration which blocked neuromuscular transmission.
A mechanism influencing the ionic flux into and out of the muscle cell in connection with excitation is discussed as a possible site of action of veratridine.
REFERENCES
Acheson, G. H., and Rosenblueth, A.: Am. J. Physiol., 133: 736, 1941.
Ahora, R. B.: This Journal, 108 : 26, 1953.
Arora, R. B., and Krayer, O.: This Journal, 106: 37,1952.
Benforado, J. M., and Witt, P. N.: This Journal, 110: 334,1954. de Fonbrune, P.: Monographies de PInstitut Pasteur: Paris, 1949.
Fuentes, J.: This Journal, 119 : 225,1957.
Graham, H., and Gasser, H.: This Journal, 43: 163, 1931.
Grupp, M.: Arch. f. exper. Path. u. Pharmakol., 212: 221, 1951.
Guttman, R.: J. Gen. Physiol., 23: 343, 1940.
Hodler, J., Staempfli, R., and Tasaki, I.: Helv. Physiol. Acta, 8: C 62, 1950.
Krayer, O., and Acheson, G. H.: Physiol. Rev., 26: 381, 1946.
Krayer, O., and George, H. W.: This Journal, 103: 249, 1951.
Ling, G., and Gerard, R. W.: J. Cell. Comp. Physiol., 34: 383, 1949.
Miyake, Y.: Arb. a. d. Med. Fakultaet, Okayama, 6: 237, 1939.
Nastuk, W. L., and Hodgkin, A. L.: J. Cell. Comp. Physiol., 35: 39, 1950.
Patón, W. D. M., and Zaimis, E.: Pharmacol. Rev., 4: 219, 1952.
Robb, J. S., and Mallov, S.: This Journal, 108 : 258, 1953.
Rosenblueth, A., Wills, G., and Hoagland, H.: Am. J. Physiol., 133: 724, 1941. Sandow, A.: The Yale J. Biol. & Med., 25: 176, 1952.
Sandow, A., and Kiebel, G.: Am. J. Physiol., 169 : 649, 1952.
Schaefer, H., and Goepfert, H.: Pflüg. Arch. ges. Physiol., 238: 684, 1937.
Schatzmann, H. J.: Helv. Physiol. Acta, 11: 346, 1953.
Schatzmann, H. J., and Witt, P. N.: This Journal, 112: 501, 1954.
Schmitt, F. O., and Gasser, H.: Am. J. Physiol., 104: 320, 1933.
Straub, R.: Helv. Physiol. Acta, 14: 1, 1956.


